

Epigenetic control of female puberty
- PMID: 23354331
- PMCID: PMC3581714
- DOI: 10.1038/nn.3319
Epigenetic control of female puberty
Abstract
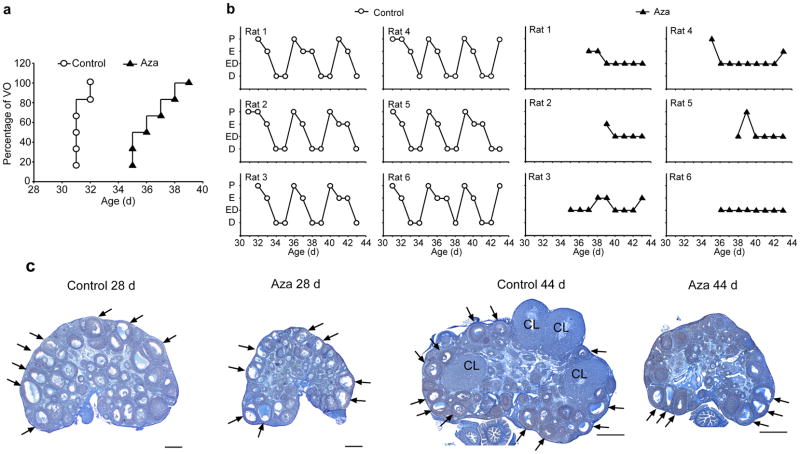
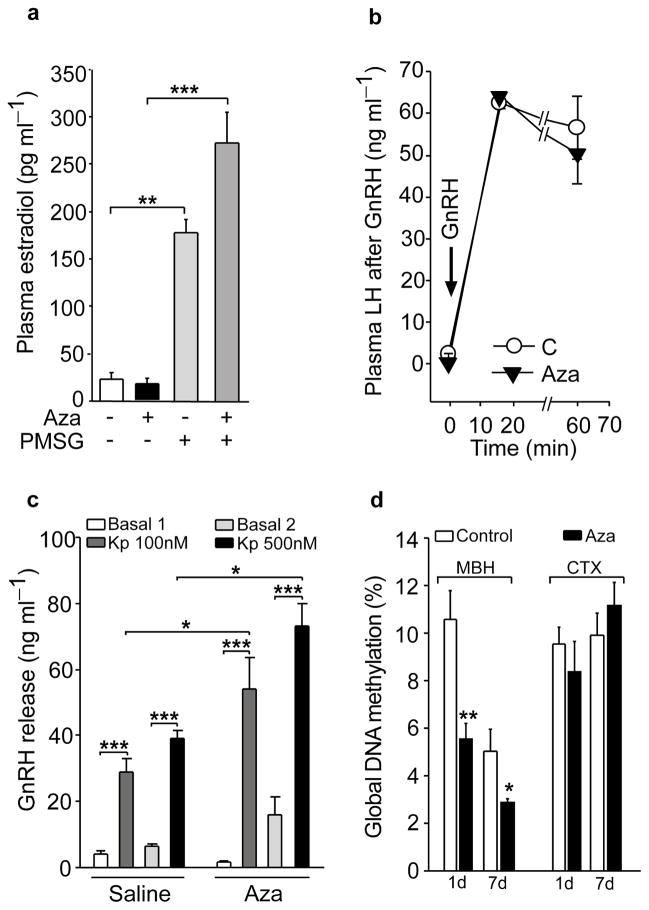
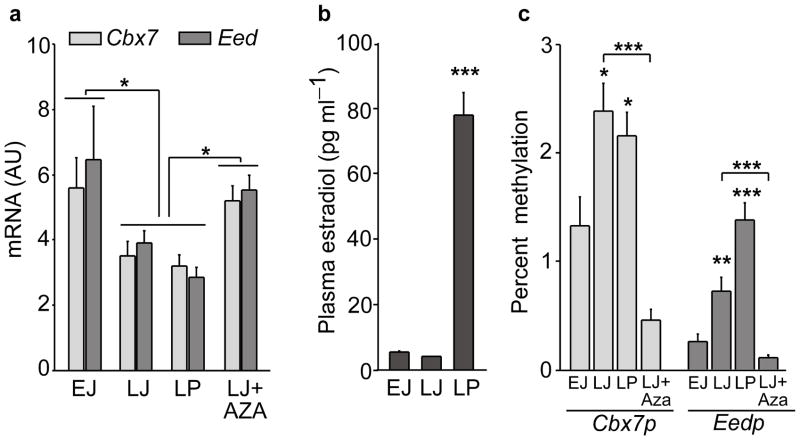
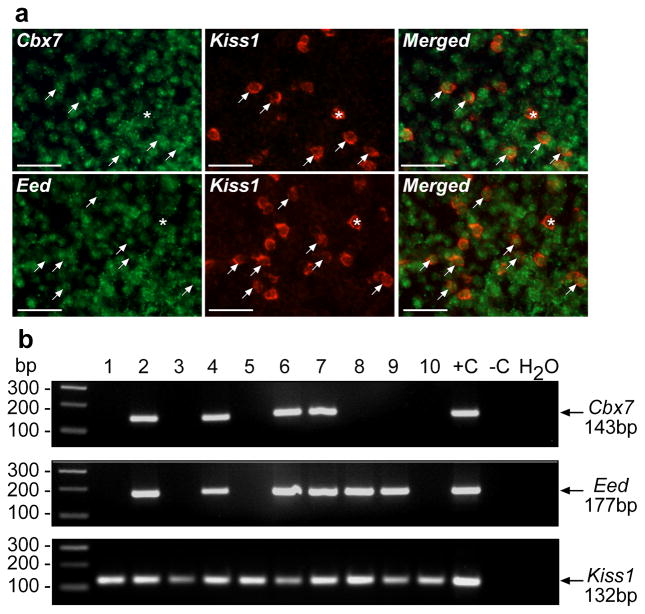
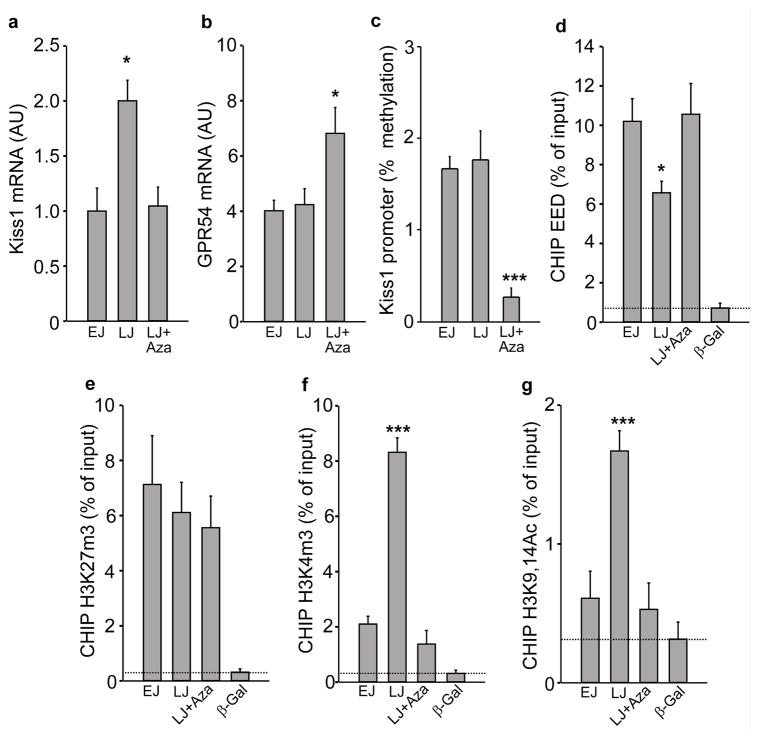
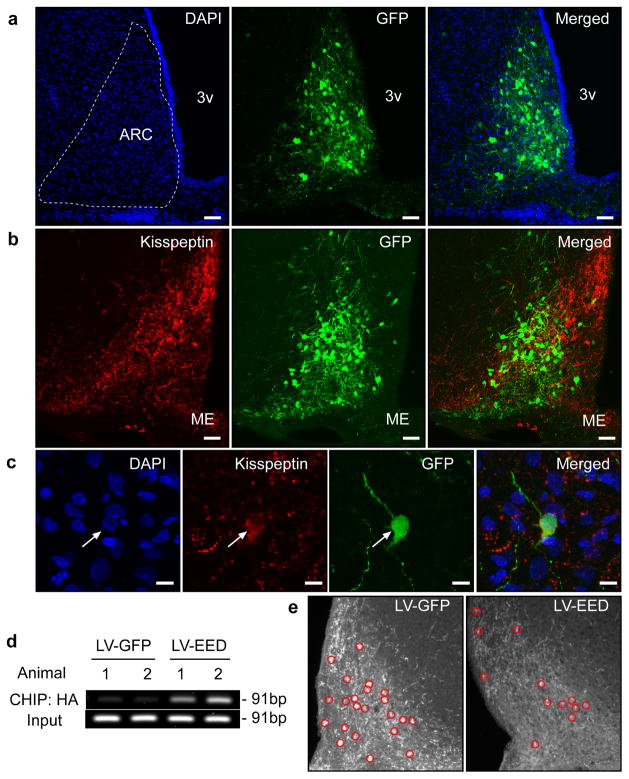
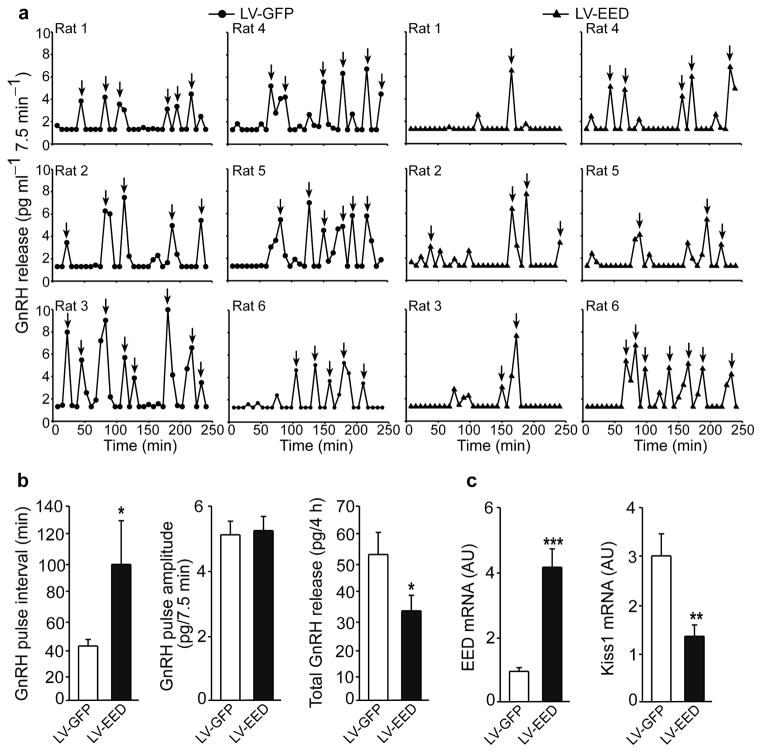
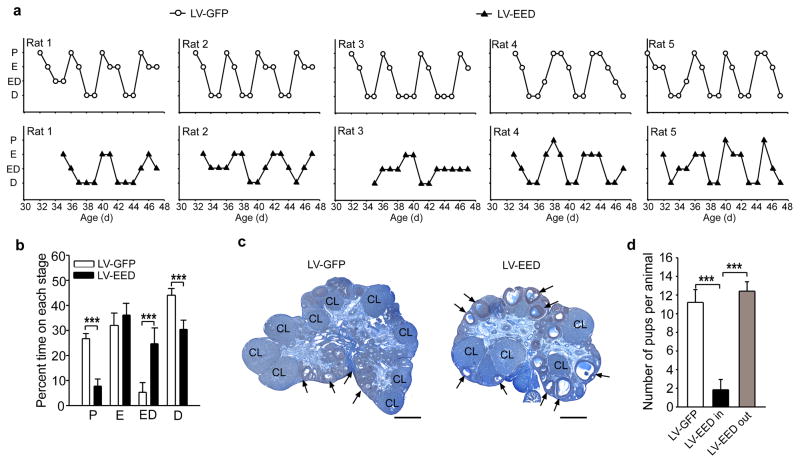
The timing of puberty is controlled by many genes. The elements coordinating this process have not, however, been identified. Here we show that an epigenetic mechanism of transcriptional repression times the initiation of female puberty in rats. We identify silencers of the Polycomb group (PcG) as principal contributors to this mechanism and show that PcG proteins repress Kiss1, a puberty-activating gene. Hypothalamic expression of two key PcG genes, Eed and Cbx7, decreased and methylation of their promoters increased before puberty. Inhibiting DNA methylation blocked both events and resulted in pubertal failure. The pubertal increase in Kiss1 expression was accompanied by EED loss from the Kiss1 promoter and enrichment of histone H3 modifications associated with gene activation. Preventing the eviction of EED from the Kiss1 promoter disrupted pulsatile gonadotropin-releasing hormone release, delayed puberty and compromised fecundity. Our results identify epigenetic silencing as a mechanism underlying the neuroendocrine control of female puberty.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Comment in
-
A piece in the puzzle of puberty.Nat Neurosci. 2013 Mar;16(3):251-3. doi: 10.1038/nn.3339. Nat Neurosci. 2013. PMID: 23434974 No abstract available.
References
-
- Plant TM, Witchel SF. Puberty in nonhuman primates and humans. In: Neill JD, editor. The Physiology of Reproduction. 3. Academic Press/Elsevier; San Diego: 2006. pp. 2177–2230.
-
- Ojeda SR, Skinner MK. Puberty in the rat. In: Neill JD, editor. The Physiology of Reproduction. 3. Academic Press/Elsevier; San Diego: 2006. pp. 2061–2126.
-
- Kordon C, Drouva SV, Martínez de la Escalera G, Weiner RI. Role of classic and peptide neuromediators in the neuroendocrine regulation of luteinizing hormone and prolactin. In: Knobil E, Neill JD, editors. The Physiology of Reproduction. 2. Vol. 1. Raven Press; New York: 1994. pp. 1621–1681.
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
