

A red/far-red light-responsive bi-stable toggle switch to control gene expression in mammalian cells
- PMID: 23355611
- PMCID: PMC3627562
- DOI: 10.1093/nar/gkt002
A red/far-red light-responsive bi-stable toggle switch to control gene expression in mammalian cells
Abstract
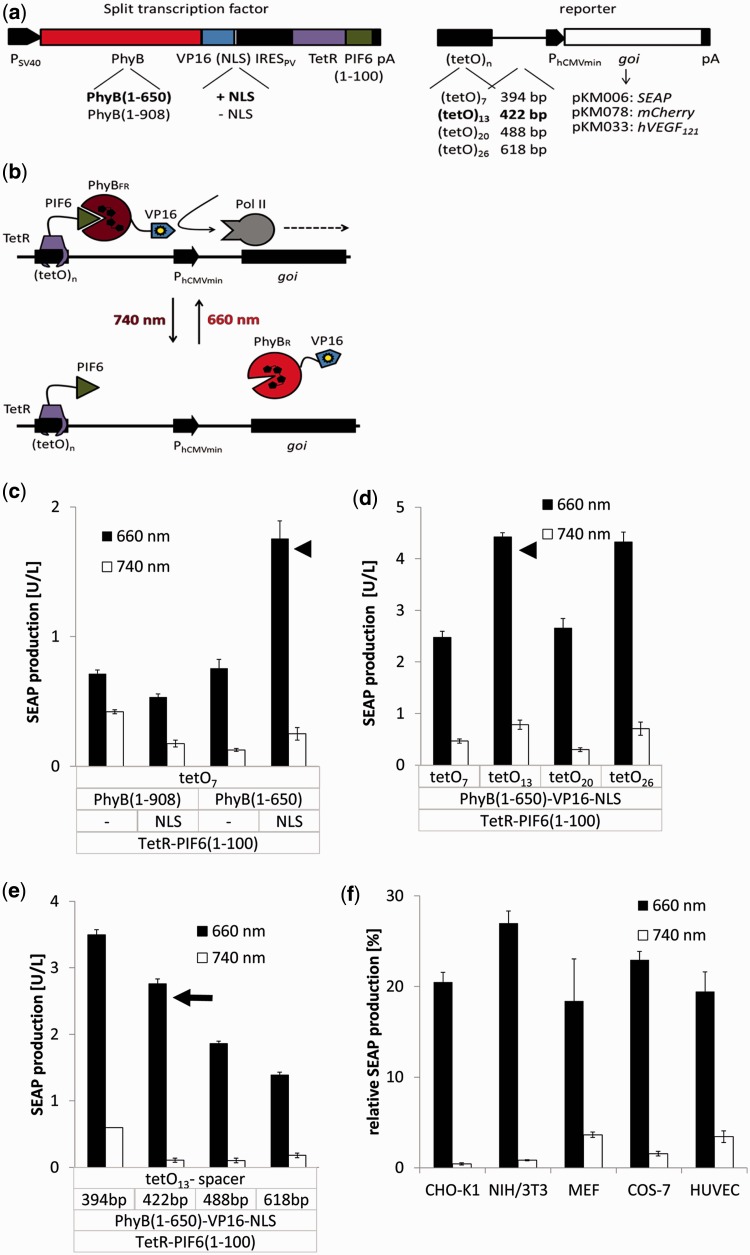
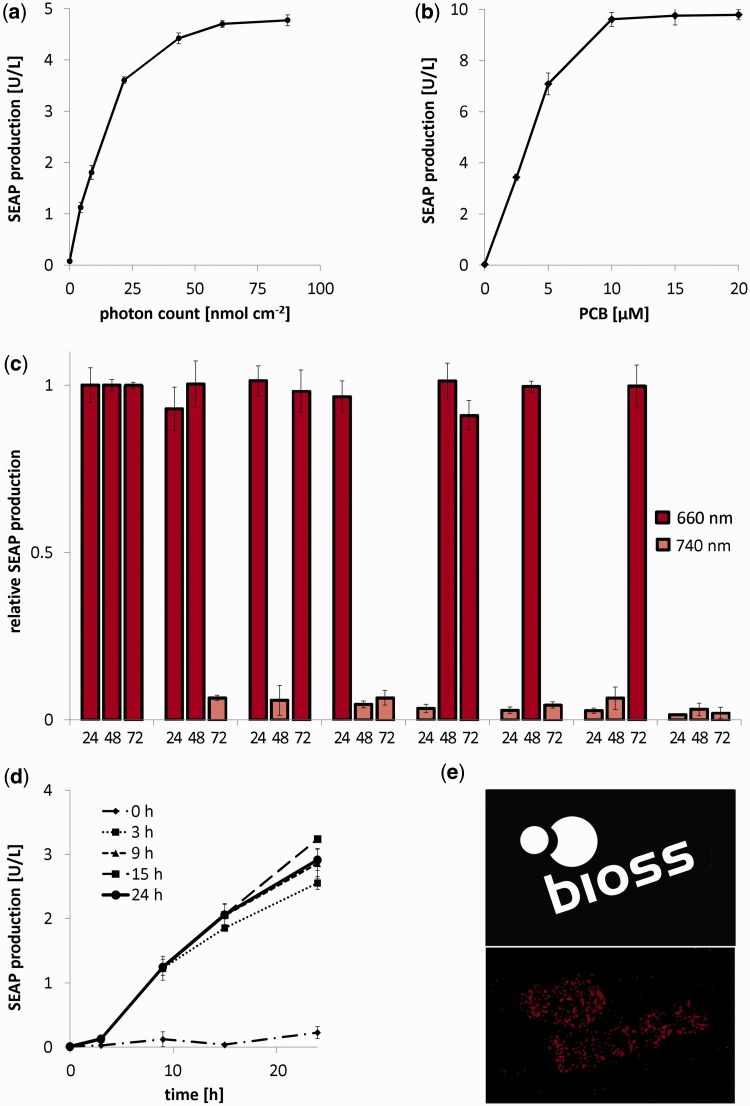
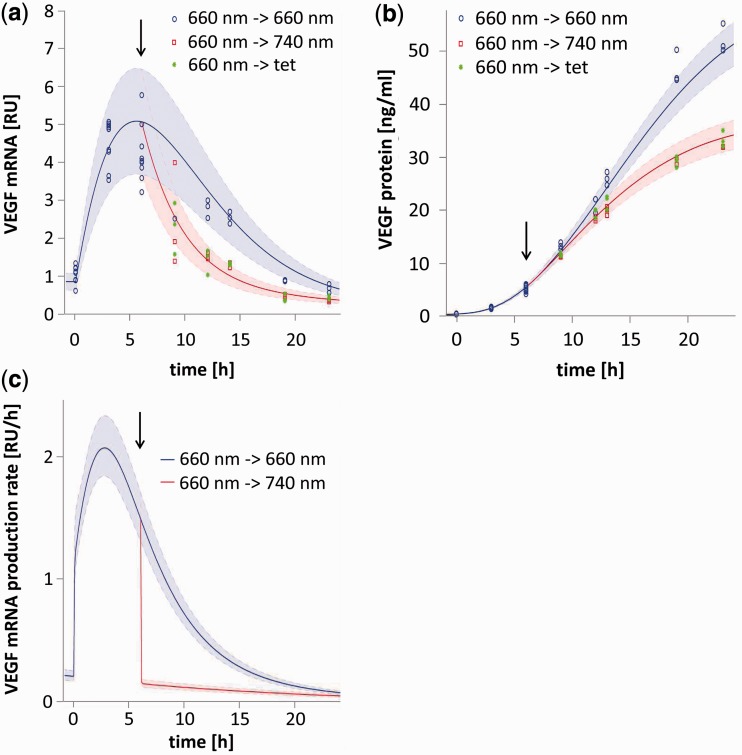
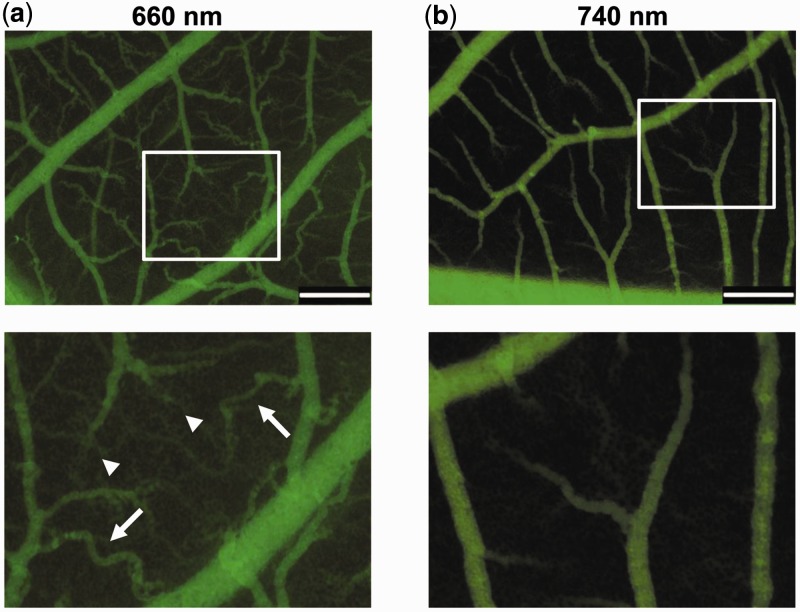
Growth and differentiation of multicellular systems is orchestrated by spatially restricted gene expression programs in specialized subpopulations. The targeted manipulation of such processes by synthetic tools with high-spatiotemporal resolution could, therefore, enable a deepened understanding of developmental processes and open new opportunities in tissue engineering. Here, we describe the first red/far-red light-triggered gene switch for mammalian cells for achieving gene expression control in time and space. We show that the system can reversibly be toggled between stable on- and off-states using short light pulses at 660 or 740 nm. Red light-induced gene expression was shown to correlate with the applied photon number and was compatible with different mammalian cell lines, including human primary cells. The light-induced expression kinetics were quantitatively analyzed by a mathematical model. We apply the system for the spatially controlled engineering of angiogenesis in chicken embryos. The system's performance combined with cell- and tissue-compatible regulating red light will enable unprecedented spatiotemporally controlled molecular interventions in mammalian cells, tissues and organisms.
Figures
Similar articles
-
Controlling gene expression with light: a multidisciplinary endeavour.Biochem Soc Trans. 2020 Aug 28;48(4):1645-1659. doi: 10.1042/BST20200014. Biochem Soc Trans. 2020. PMID: 32657338 Free PMC article. Review.
-
A red light-controlled synthetic gene expression switch for plant systems.Mol Biosyst. 2014 Jul;10(7):1679-88. doi: 10.1039/c3mb70579j. Epub 2014 Jan 27. Mol Biosyst. 2014. PMID: 24469598
-
Control of gene expression using a red- and far-red light-responsive bi-stable toggle switch.Nat Protoc. 2014 Mar;9(3):622-32. doi: 10.1038/nprot.2014.038. Epub 2014 Feb 20. Nat Protoc. 2014. PMID: 24556785
-
Optogenetics in Plants: Red/Far-Red Light Control of Gene Expression.Methods Mol Biol. 2016;1408:125-39. doi: 10.1007/978-1-4939-3512-3_9. Methods Mol Biol. 2016. PMID: 26965120
-
Phytochrome-interacting factor from Arabidopsis to liverwort.Curr Opin Plant Biol. 2017 Feb;35:54-60. doi: 10.1016/j.pbi.2016.11.004. Epub 2016 Nov 19. Curr Opin Plant Biol. 2017. PMID: 27875778 Review.
Cited by
-
Controlling gene expression with light: a multidisciplinary endeavour.Biochem Soc Trans. 2020 Aug 28;48(4):1645-1659. doi: 10.1042/BST20200014. Biochem Soc Trans. 2020. PMID: 32657338 Free PMC article. Review.
-
Algal photoreceptors: in vivo functions and potential applications.Planta. 2014 Jan;239(1):1-26. doi: 10.1007/s00425-013-1962-5. Epub 2013 Oct 1. Planta. 2014. PMID: 24081482 Review.
-
Engineering light-controllable CAR T cells for cancer immunotherapy.Sci Adv. 2020 Feb 19;6(8):eaay9209. doi: 10.1126/sciadv.aay9209. eCollection 2020 Feb. Sci Adv. 2020. PMID: 32128416 Free PMC article.
-
Tools for controlling protein interactions using light.Curr Protoc Cell Biol. 2014 Sep 2;64:17.16.1-20. doi: 10.1002/0471143030.cb1716s64. Curr Protoc Cell Biol. 2014. PMID: 25181301 Free PMC article. Review.
-
Synthetic dual-input mammalian genetic circuits enable tunable and stringent transcription control by chemical and light.Nucleic Acids Res. 2016 Apr 7;44(6):2677-90. doi: 10.1093/nar/gkv1343. Epub 2015 Dec 15. Nucleic Acids Res. 2016. PMID: 26673714 Free PMC article.
References
-
- Wang X, Chen X, Yang Y. Spatiotemporal control of gene expression by a light-switchable transgene system. Nat. Methods. 2012;9:266–269. - PubMed
-
- Yazawa M, Sadaghiani AM, Hsueh B, Dolmetsch RE. Induction of protein-protein interactions in live cells using light. Nat. Biotechnol. 2009;27:941–945. - PubMed
-
- Ye HF, Daoud-El Baba M, Peng RW, Fussenegger M. A synthetic optogenetic transcription device enhances blood-glucose homeostasis in mice. Science. 2011;332:1565–1568. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
