

The energy and work of a ligand-gated ion channel
- PMID: 23357172
- PMCID: PMC4407511
- DOI: 10.1016/j.jmb.2013.01.027
The energy and work of a ligand-gated ion channel
Abstract
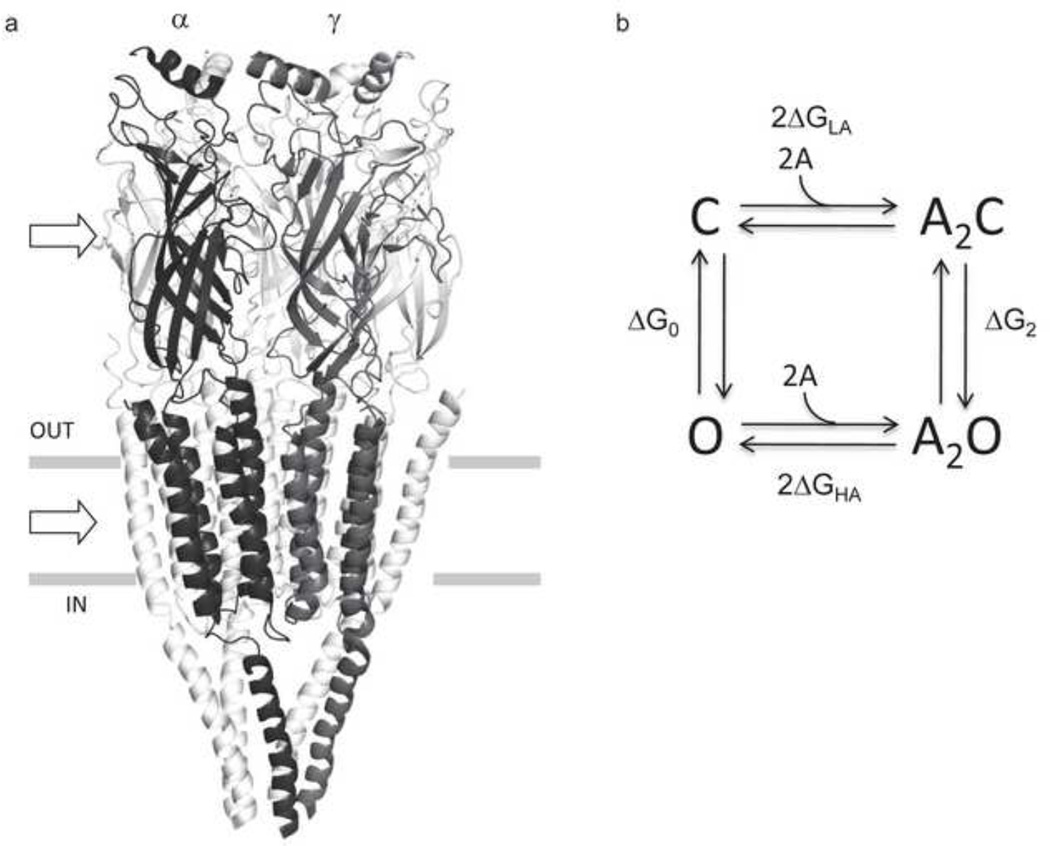
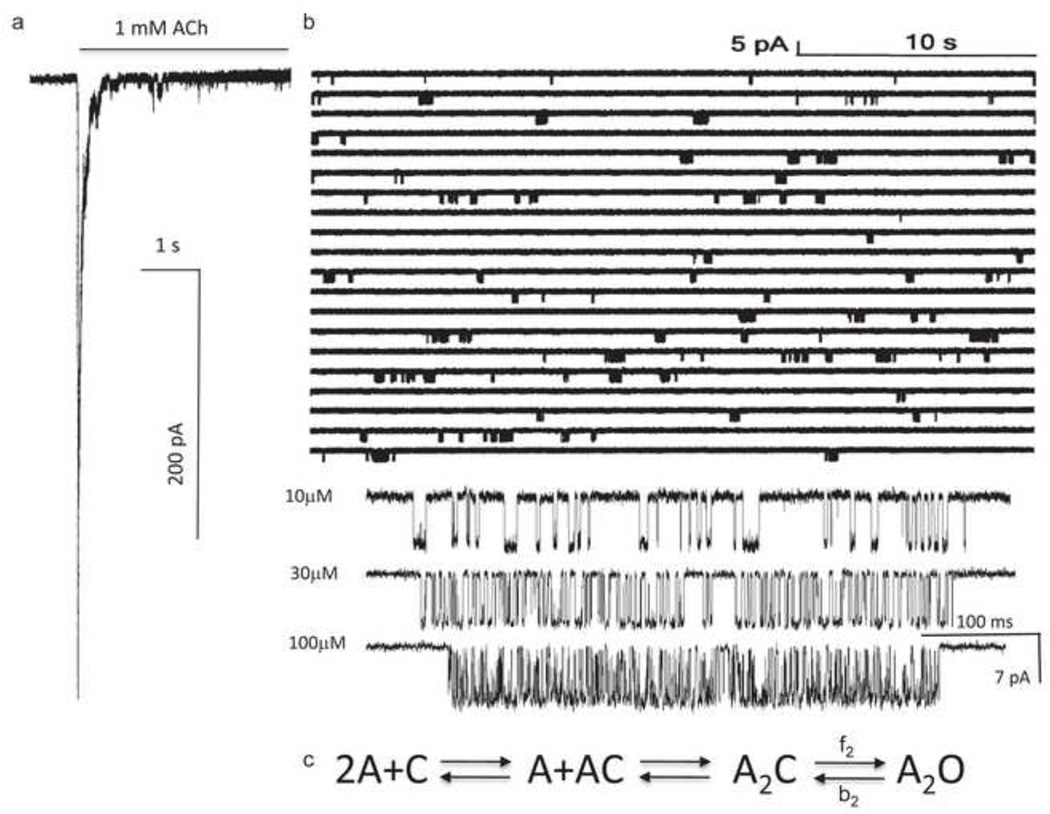
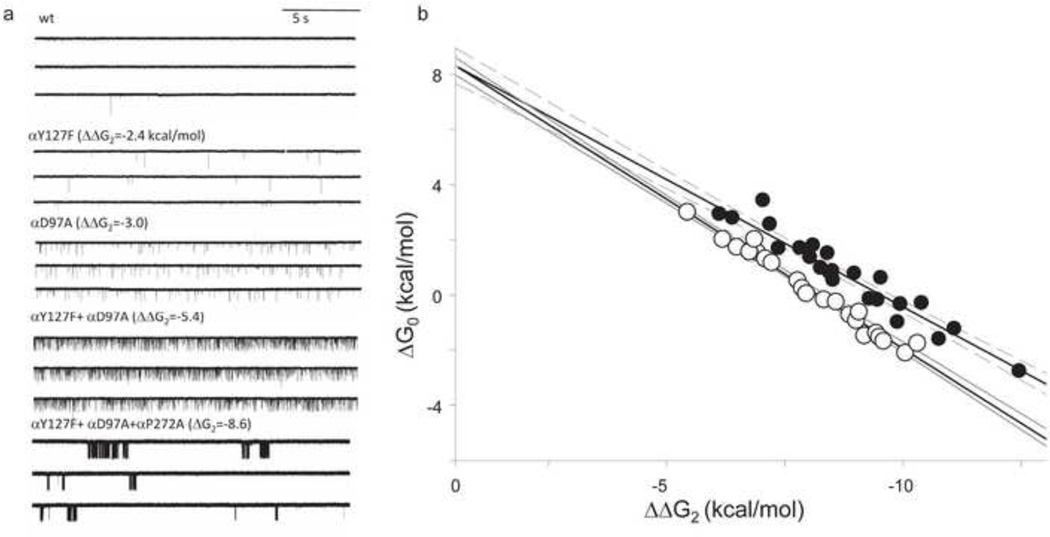
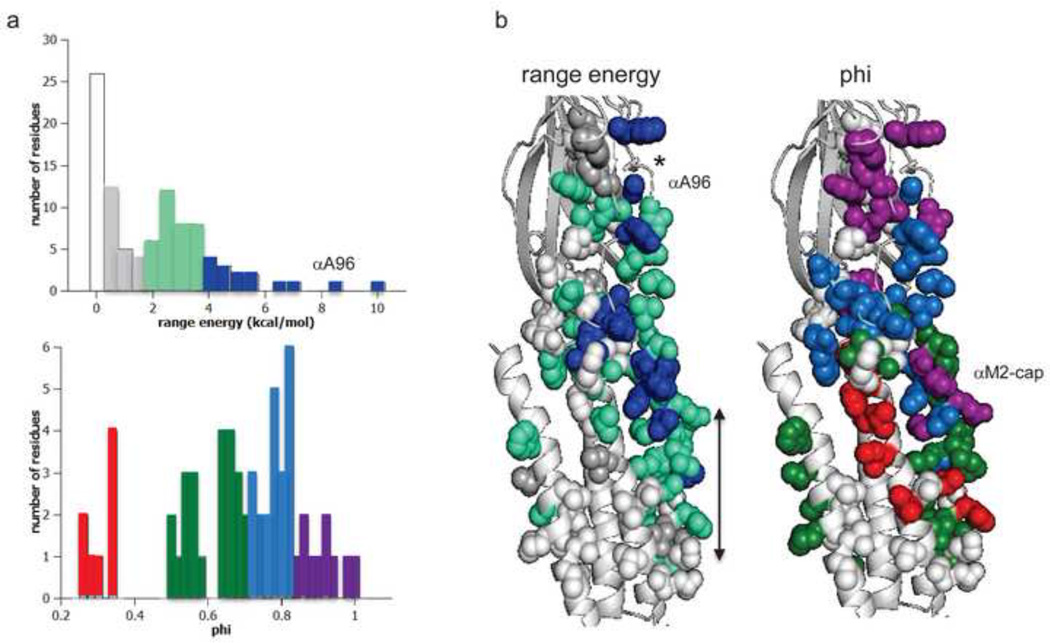
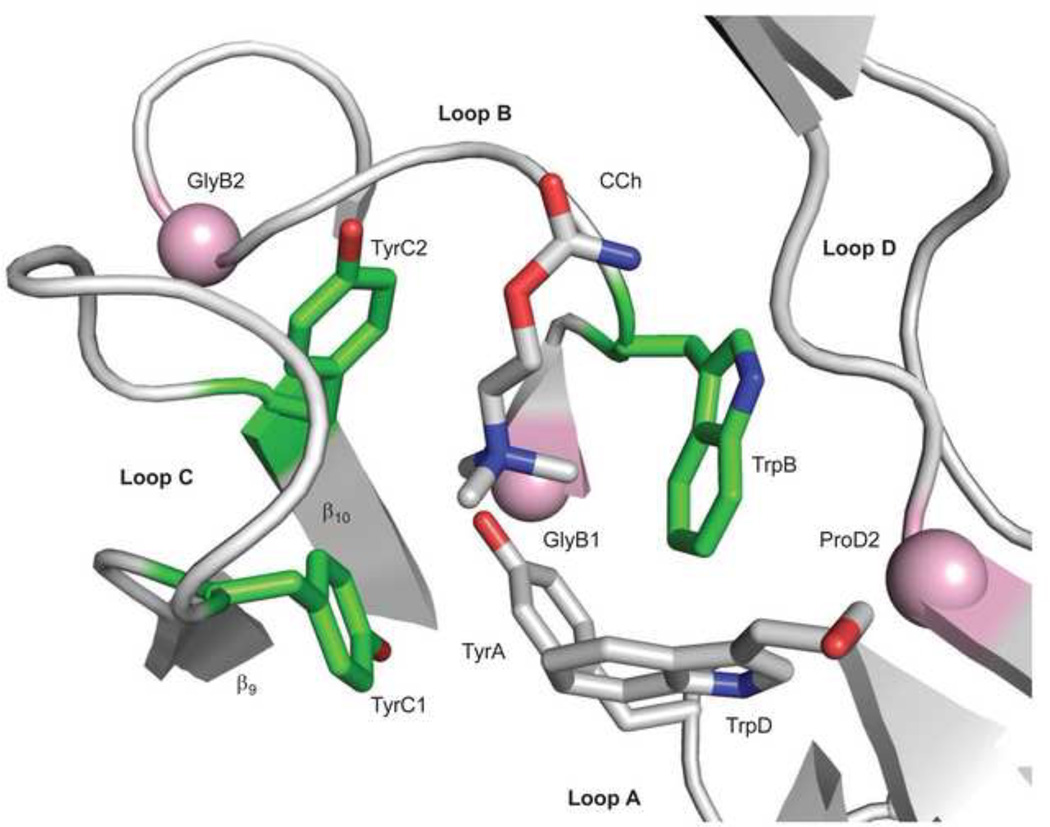
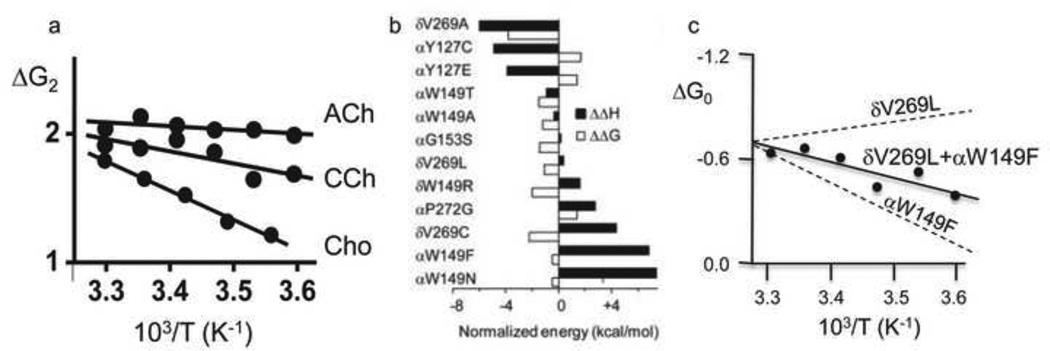
Ligand-gated ion channels are allosteric membrane proteins that isomerize between C(losed) and O(pen) conformations. A difference in affinity for ligands in the two states influences the C↔O "gating" equilibrium constant. The energies associated with adult-type mouse neuromuscular nicotinic acetylcholine receptor (AChR) channel gating have been measured by using single-channel electrophysiology. Without ligands, the free energy, enthalpy and entropy of gating are ΔG0=+8.4, ΔH0=+10.9 and TΔS0=+2.5kcal/mol (-100mV, 23°C). Many mutations throughout the protein change ΔG0, including natural ones that cause disease. Agonists and most mutations change approximately independently the ground-state energy difference; thus, it is possible to forecast and engineer AChR responses simply by combining perturbations. The free energy of the low↔high affinity change for the neurotransmitter at each of two functionally equivalent binding sites is ΔGB(ACh)=-5.1kcal/mol. ΔGB(ACh) is set mainly by interactions of ACh with just three binding site aromatic groups. For a series of structurally related agonists, there is a correlation between the energies of low- and high-affinity binding, which implies that gating commences with the formation of the low-affinity complex. Brief, intermediate states in binding and gating have been detected. Several proposals for the nature of the gating transition-state energy landscape and the isomerization mechanism are discussed.
Copyright © 2013. Published by Elsevier Ltd.
Figures
References
-
- Corringer P-J, Poitevin F, Prevost, Marie S, Sauguet L, Delarue M, Changeux J-P. Structure and Pharmacology of Pentameric Receptor Channels: From Bacteria to Brain. Structure. 2012;20:941–956. - PubMed
-
- Monod J, Wyman J, Changeux JP. On the Nature of Allosteric Transitions: A Plausible Model. J Mol Biol. 1965;12:88–118. - PubMed
-
- Karlin A. On the application of "a plausible model" of allosteric proteins to the receptor for acetylcholine. J Theor Biol. 1967;16:306–320. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
