

Imbalance of desmoplastic stromal cell numbers drives aggressive cancer processes
- PMID: 23359139
- PMCID: PMC4034674
- DOI: 10.1002/path.4172
Imbalance of desmoplastic stromal cell numbers drives aggressive cancer processes
Abstract
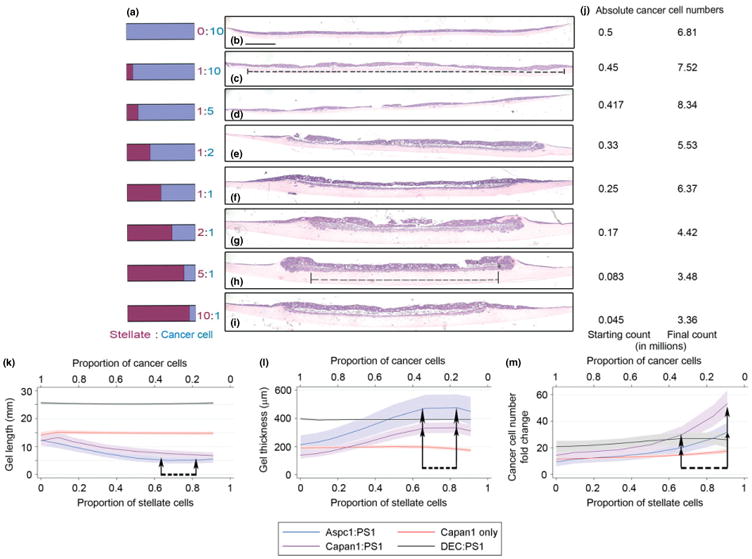
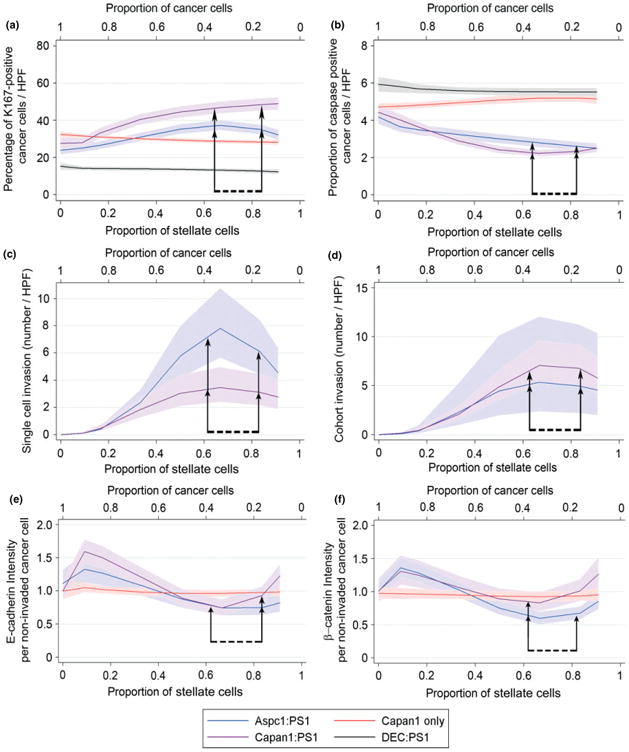
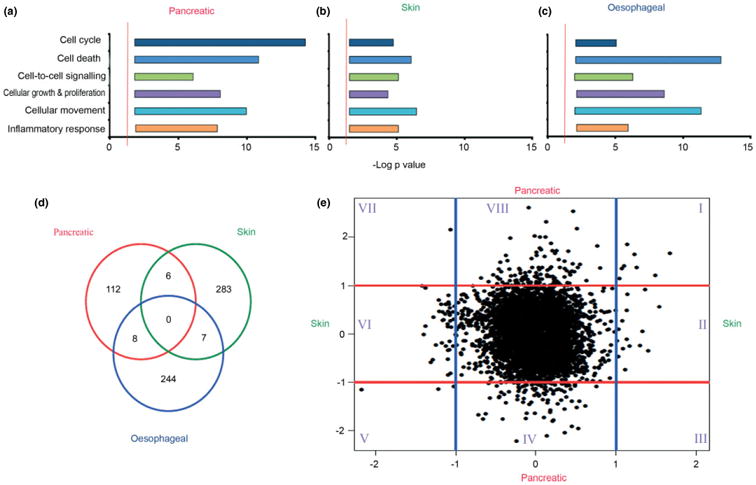
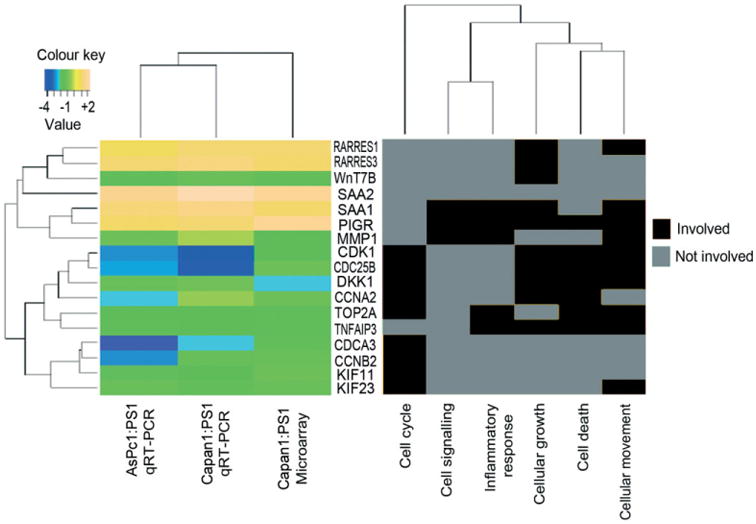
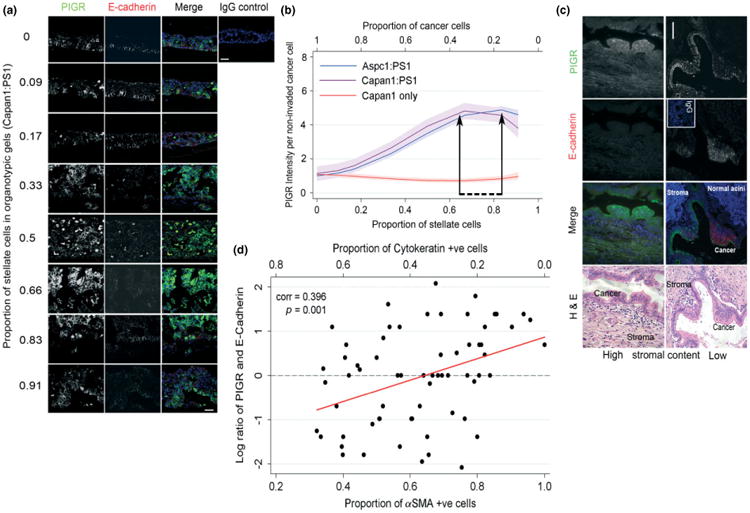
Epithelial tissues have sparse stroma, in contrast to their corresponding tumours. The effect of cancer cells on stromal cells is well recognized. Increasingly, stromal components, such as endothelial and immune cells, are considered indispensable for cancer progression. The role of desmoplastic stroma, in contrast, is poorly understood. Targeting such cellular components within the tumour is attractive. Recent evidence strongly points towards a dynamic stromal cell participation in cancer progression that impacts patient prognosis. The role of specific desmoplastic stromal cells, such as stellate cells and myofibroblasts in pancreatic, oesophageal and skin cancers, was studied in bio-engineered, physiomimetic organotypic cultures and by regression analysis. For pancreatic cancer, the maximal effect on increasing cancer cell proliferation and invasion, as well as decreasing cancer cell apoptosis, occurs when stromal (pancreatic stellate cells) cells constitute the majority of the cellular population (maximal effect at a stromal cell proportion of 0.66-0.83), accompanied by change in expression of key molecules such as E-cadherin and β-catenin. Gene-expression microarrays, across three tumour types, indicate that stromal cells consistently and significantly alter global cancer cell functions such as cell cycle, cell-cell signalling, cell movement, cell death and inflammatory response. However, these changes are mediated through cancer type-specific alteration of expression, with very few common targets across tumour types. As highlighted by these in vitro data, the reciprocal relationship of E-cadherin and polymeric immunoglobulin receptor (PIGR) expression in cancer cells could be shown, in vivo, to be dependent on the stromal content of human pancreatic cancer. These studies demonstrate that context-specific cancer-stroma crosstalk requires to be precisely defined for effective therapeutic targeting. These data may be relevant to non-malignant processes where epithelial cells interact with stromal cells, such as chronic inflammatory and fibrotic conditions.
Copyright © 2013 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
Conflict of interest statement
No conflicts of interest were declared.
Figures
Comment in
-
Understanding the stroma of pancreatic cancer: co-evolution of the microenvironment with epithelial carcinogenesis.J Pathol. 2013 Sep;231(1):4-7. doi: 10.1002/path.4213. Epub 2013 Jul 8. J Pathol. 2013. PMID: 23716361
References
-
- Erkan M, Michalski CW, Rieder S, et al. The activated stroma index is a novel and independent prognostic marker in pancreatic ductal adenocarcinoma. Clin Gastroenterol Hepatol. 2008;6:1155–1161. - PubMed
-
- Fujita H, Ohuchida K, Mizumoto K, et al. α-Smooth muscle actin expressing stroma promotes an aggressive tumor biology in pancreatic ductal adenocarcinoma. Pancreas. 2010;39:1254–1262. - PubMed
-
- Wang K, Ma W, Wang J, et al. Tumor:stroma ratio is an independent predictor for survival in esophageal squamous cell carcinoma. J Thorac Oncol. 2012;7:1457–1461. - PubMed
-
- Woenne EC, Lederle W, Zwick S, et al. MMP inhibition blocks fibroblast-dependent skin cancer invasion, reduces vascularization and alters VEGF-A and PDGF-BB expression. Anticancer Res. 2012;30:703–711. - PubMed
-
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–674. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
