

The small GTPase Rif is dispensable for platelet filopodia generation in mice
- PMID: 23359340
- PMCID: PMC3554654
- DOI: 10.1371/journal.pone.0054663
The small GTPase Rif is dispensable for platelet filopodia generation in mice
Abstract
Background: Formation of filopodia and other shape change events are vital for platelet hemostatic function. The mechanisms regulating filopodia formation by platelets are incompletely understood however. In particular the small GTPase responsible for initiating filopodia formation by platelets remains elusive. The canonical pathway involving Cdc42 is not essential for filopodia formation in mouse platelets. The small GTPase Rif (RhoF) provides an alternative route to filopodia generation in other cell types and is expressed in both human and mouse platelets.
Hypothesis/objective: We hypothesized that Rif might be responsible for generating filopodia by platelets and generated a novel knockout mouse model to investigate the functional role of Rif in platelets.
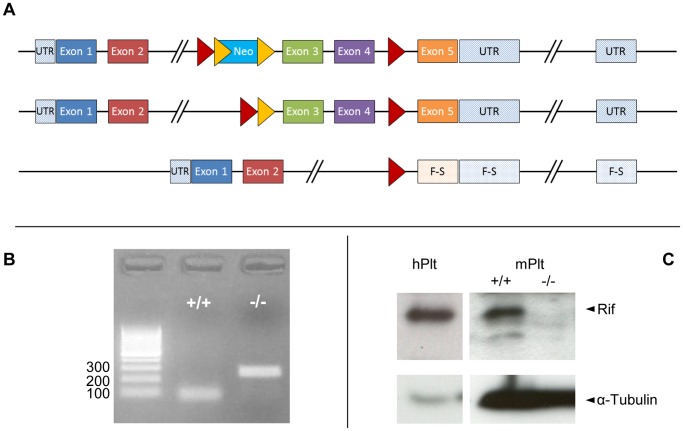
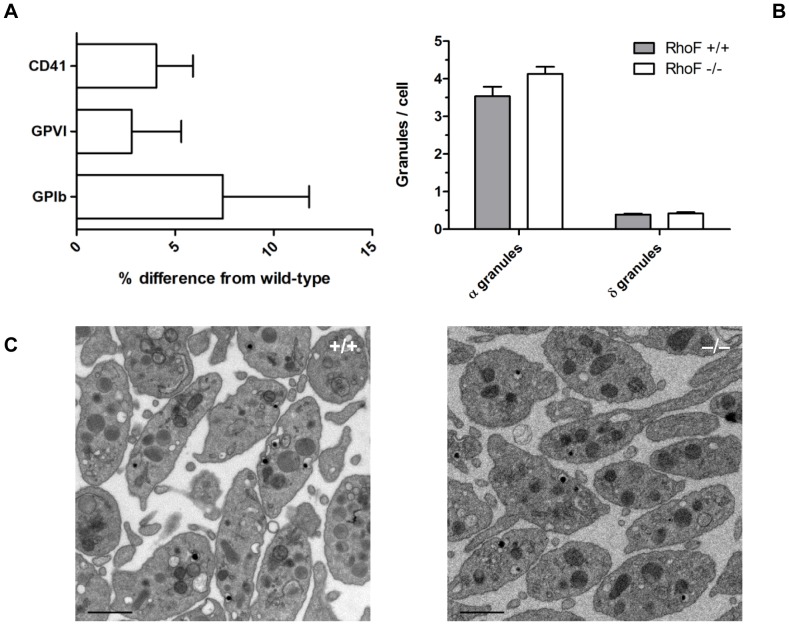
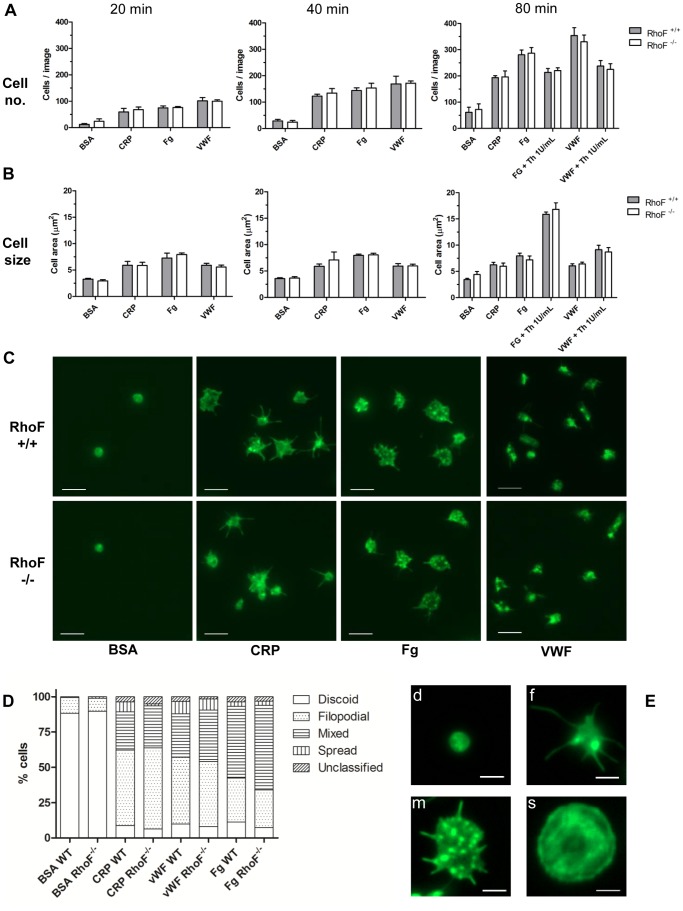
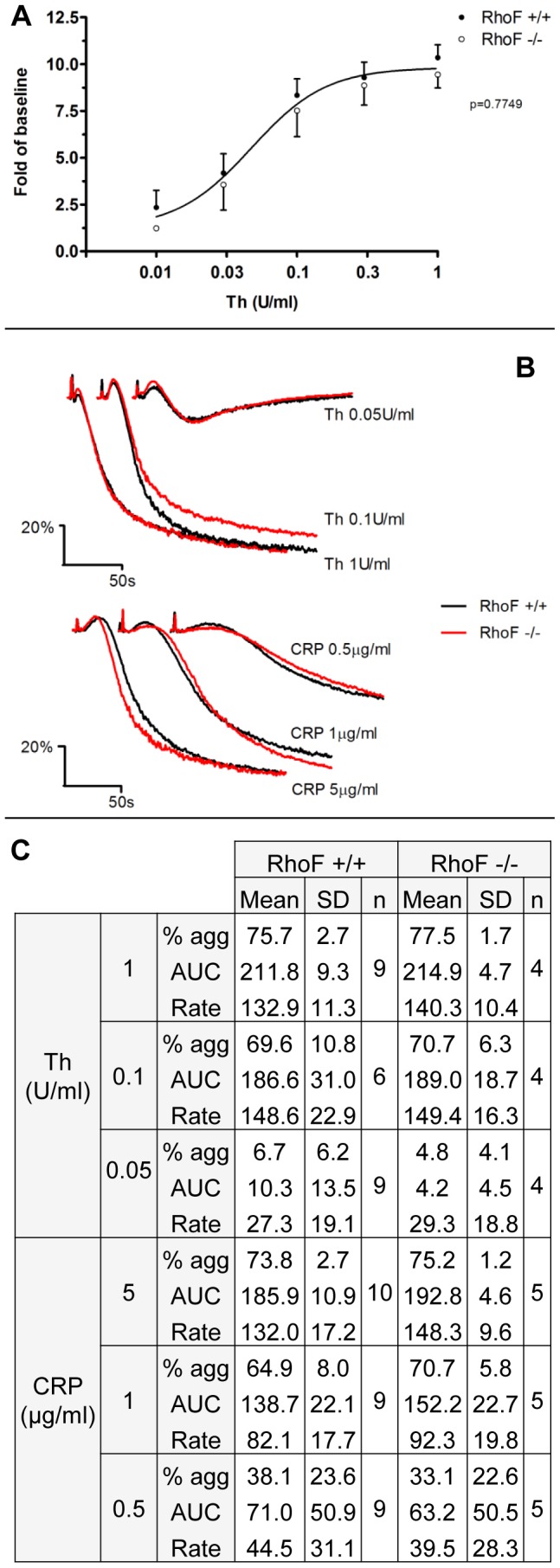
Methodology/principal findings: Constitutive RhoF(-/-) mice are viable and have normal platelet, leukocyte and erythrocyte counts and indices. RhoF(-/-) platelets form filopodia and spread normally on various agonist surfaces in static conditions and under arterial shear. In addition, RhoF(-/-) platelets have normal actin dynamics, are able to activate and aggregate normally and secrete from alpha and dense granules in response to collagen related peptide and thrombin stimulation.
Conclusions: The small GTPase Rif does not appear to be critical for platelet function in mice. Functional overlap between Rif and other small GTPases may be responsible for the non-essential role of Rif in platelets.
Conflict of interest statement
Figures
References
-
- Hartwig JH (2007) The platelet cytoskeleton. In: Michelson AD, editor. Platelets. 2 ed: Academic Press. 75–98.
-
- Eliautou S, Mangin PH, Walter U, Gachet C, Lanza F (2009) Normal filopodia extension in VASP-deficient platelets upon activation by adhesive matrices or soluble agonists. Thromb Haemost 102: 792–794. - PubMed
-
- Leng L, Kashiwagi H, Ren XD, Shattil SJ (1998) RhoA and the function of platelet integrin alphaIIbbeta3. Blood 91: 4206–4215. - PubMed
-
- Chang JC, Chang HH, Lin CT, Lo SJ (2005) The integrin alpha6beta1 modulation of PI3K and Cdc42 activities induces dynamic filopodium formation in human platelets. J Biomed Sci 12: 881–898. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
