

Prophase pathway-dependent removal of cohesin from human chromosomes requires opening of the Smc3-Scc1 gate
- PMID: 23361318
- PMCID: PMC3590994
- DOI: 10.1038/emboj.2013.7
Prophase pathway-dependent removal of cohesin from human chromosomes requires opening of the Smc3-Scc1 gate
Abstract
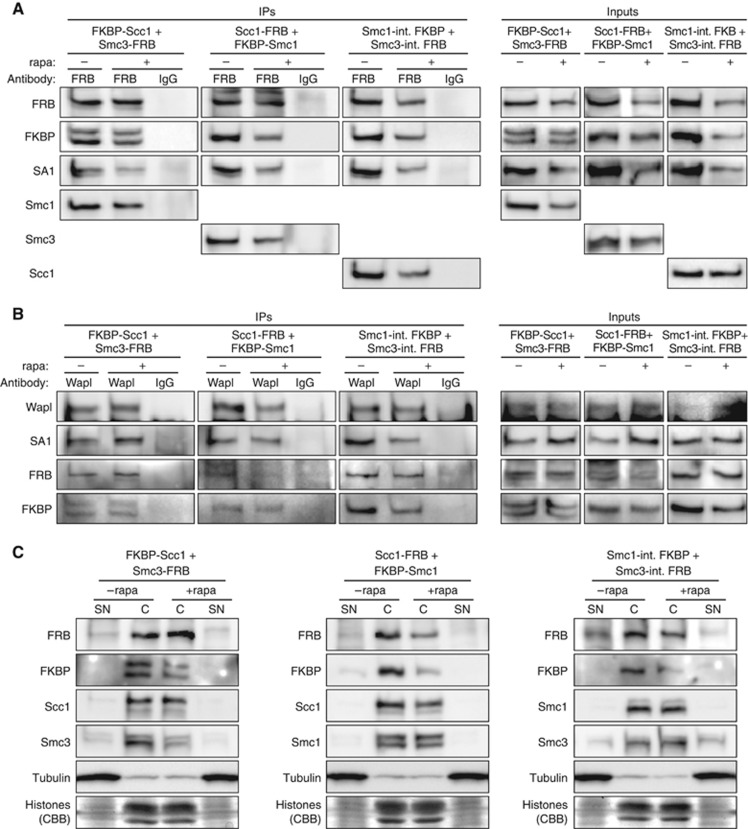
Faithful transmission of chromosomes during eukaryotic cell division requires sister chromatids to be paired from their generation in S phase until their separation in M phase. Cohesion is mediated by the cohesin complex, whose Smc1, Smc3 and Scc1 subunits form a tripartite ring that entraps both DNA double strands. Whereas centromeric cohesin is removed in late metaphase by Scc1 cleavage, metazoan cohesin at chromosome arms is displaced already in prophase by proteolysis-independent signalling. Which of the three gates is triggered by the prophase pathway to open has remained enigmatic. Here, we show that displacement of human cohesin from early mitotic chromosomes requires dissociation of Smc3 from Scc1 but no opening of the other two gates. In contrast, loading of human cohesin onto chromatin in telophase occurs through the Smc1-Smc3 hinge. We propose that the use of differently regulated gates for loading and release facilitates unidirectionality of DNA's entry into and exit from the cohesin ring.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures






References
-
- Banaszynski LA, Liu CW, Wandless TJ (2005) Characterization of the FKBP.rapamycin.FRB ternary complex. J Am Chem Soc 127: 4715–4721 - PubMed
-
- Boos D, Kuffer C, Lenobel R, Körner R, Stemmann O (2008) Phosphorylation-dependent binding of cyclin B1 to a Cdc6-like domain of human separase. J Biol Chem 283: 816–823 - PubMed
-
- Ciosk R, Shirayama M, Shevchenko A, Tanaka T, Tóth A, Shevchenko A, Nasmyth K (2000) Cohesin's binding to chromosomes depends on a separate complex consisting of Scc2 and Scc4 proteins. Mol Cell 5: 243–254 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
