

Assembly in G1 phase and long-term stability are unique intrinsic features of CENP-A nucleosomes
- PMID: 23363600
- PMCID: PMC3608502
- DOI: 10.1091/mbc.E13-01-0034
Assembly in G1 phase and long-term stability are unique intrinsic features of CENP-A nucleosomes
Abstract
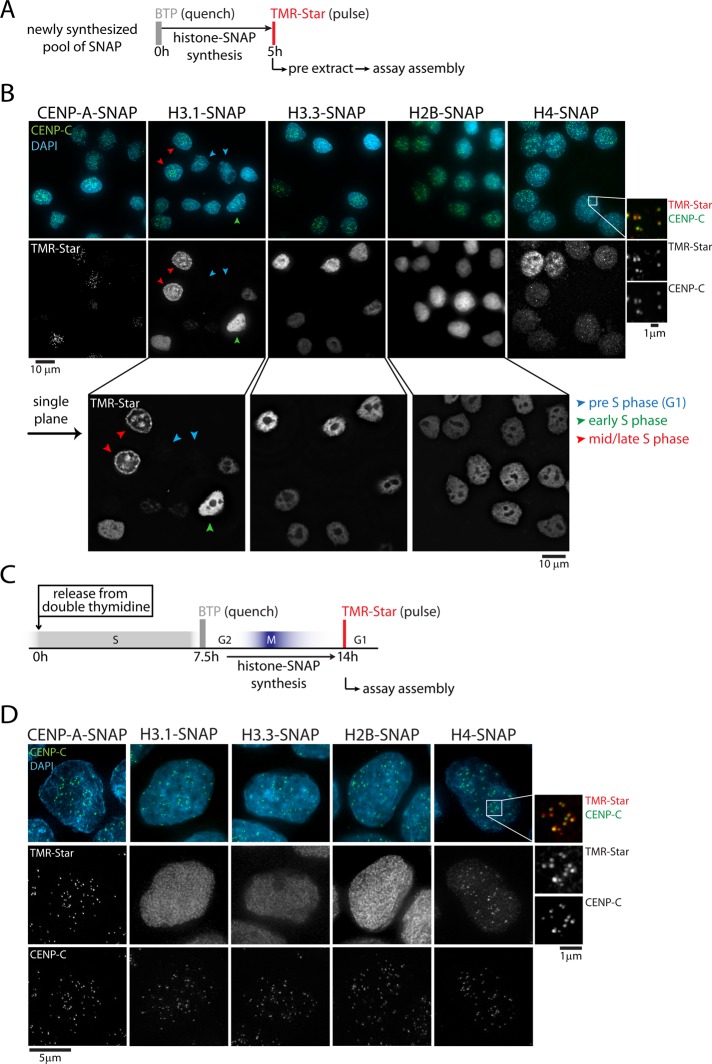
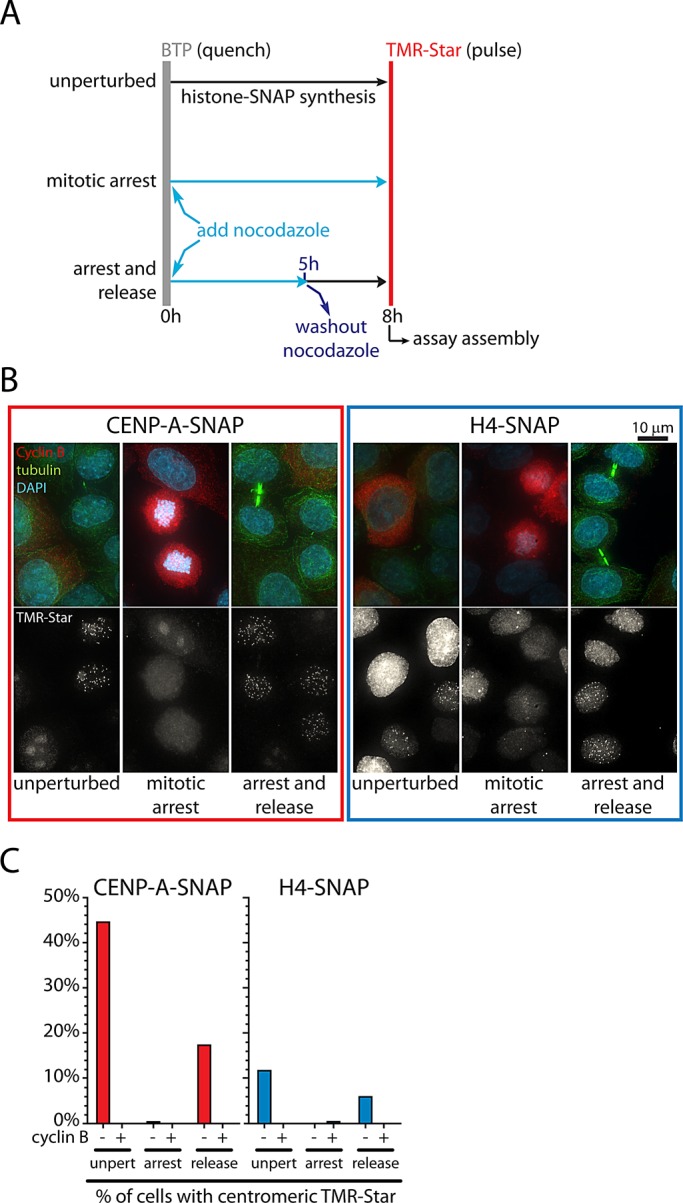
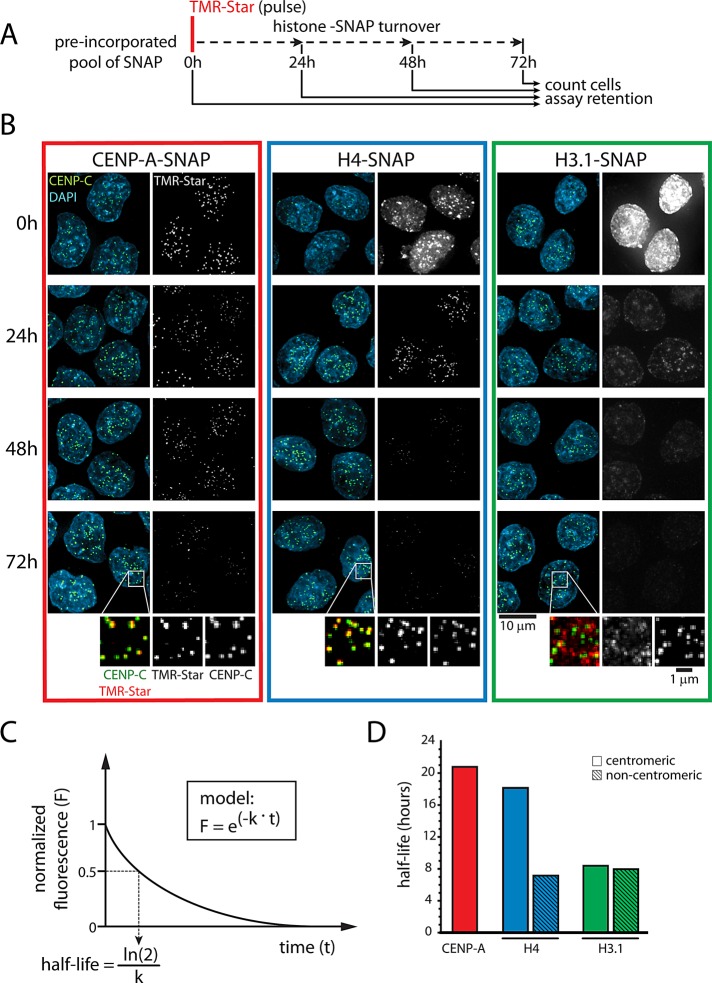
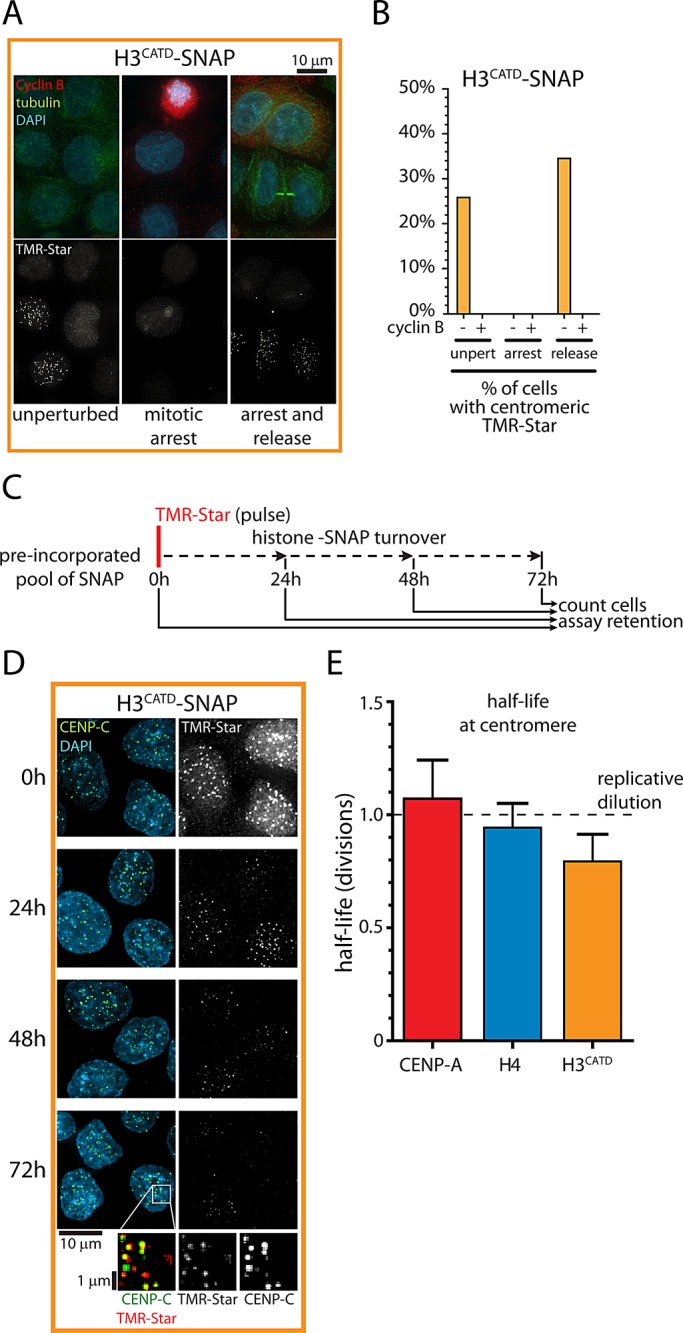
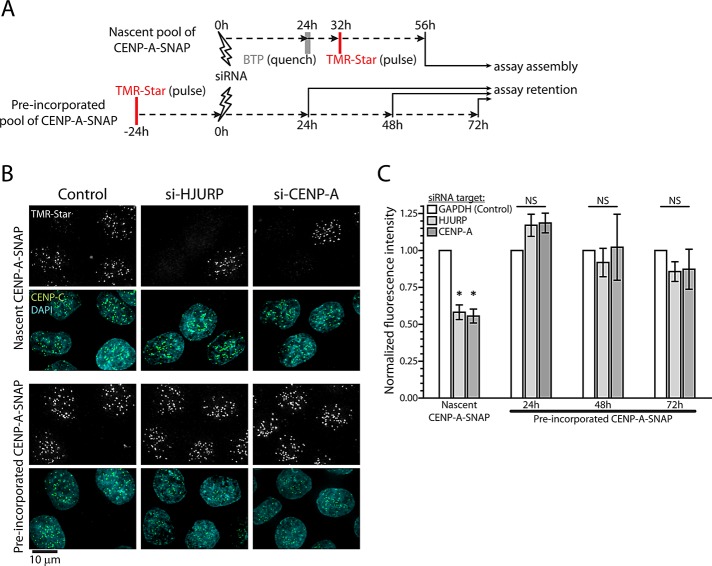
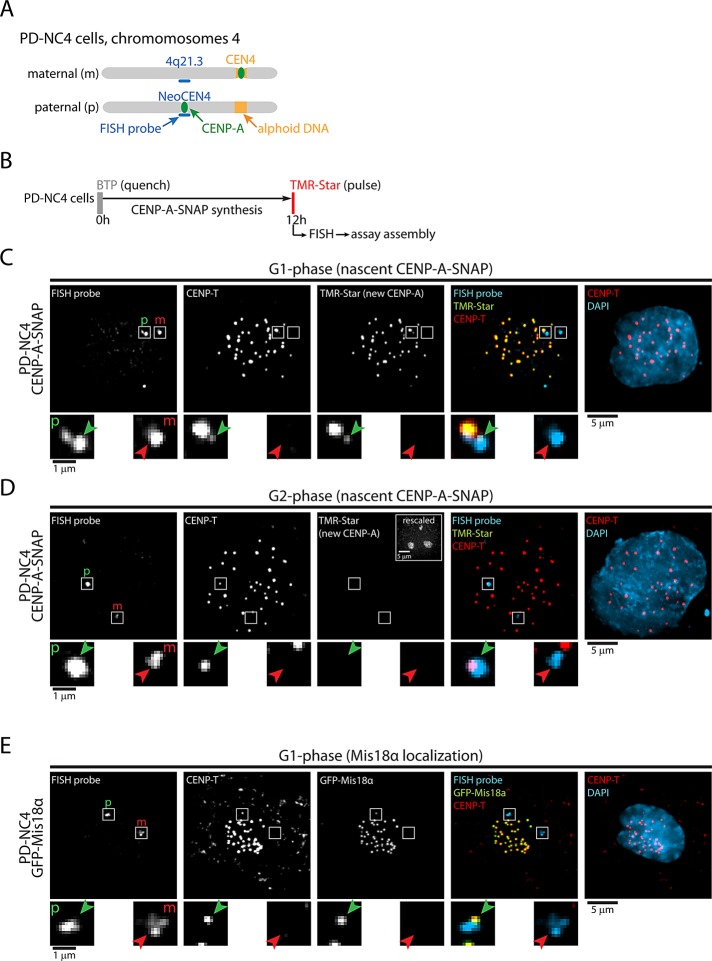
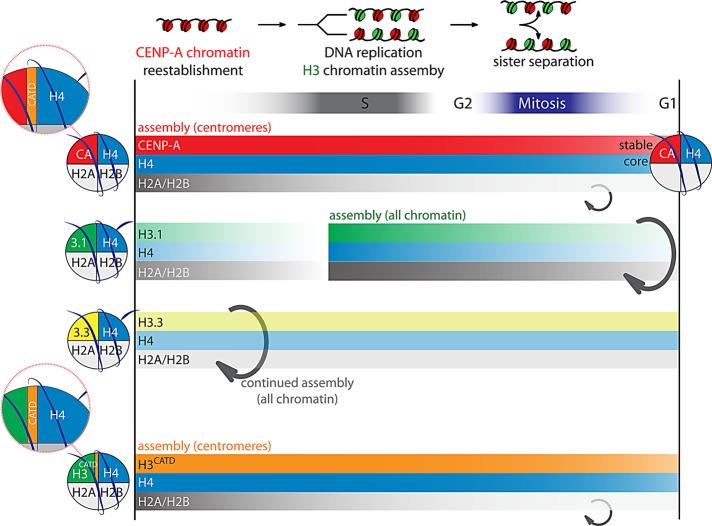
Centromeres are the site of kinetochore formation during mitosis. Centromere protein A (CENP-A), the centromere-specific histone H3 variant, is essential for the epigenetic maintenance of centromere position. Previously we showed that newly synthesized CENP-A is targeted to centromeres exclusively during early G1 phase and is subsequently maintained across mitotic divisions. Using SNAP-based fluorescent pulse labeling, we now demonstrate that cell cycle-restricted chromatin assembly at centromeres is unique to CENP-A nucleosomes and does not involve assembly of other H3 variants. Strikingly, stable retention is restricted to the CENP-A/H4 core of the nucleosome, which we find to outlast general chromatin across several cell divisions. We further show that cell cycle timing of CENP-A assembly is independent of centromeric DNA sequences and instead is mediated by the CENP-A targeting domain. Unexpectedly, this domain also induces stable transmission of centromeric nucleosomes, independent of the CENP-A deposition factor HJURP. This demonstrates that intrinsic properties of the CENP-A protein direct its cell cycle-restricted assembly and induces quantitative mitotic transmission of the CENP-A/H4 nucleosome core, ensuring long-term stability and epigenetic maintenance of centromere position.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
