

Rap1 and Canoe/afadin are essential for establishment of apical-basal polarity in the Drosophila embryo
- PMID: 23363604
- PMCID: PMC3608504
- DOI: 10.1091/mbc.E12-10-0736
Rap1 and Canoe/afadin are essential for establishment of apical-basal polarity in the Drosophila embryo
Abstract
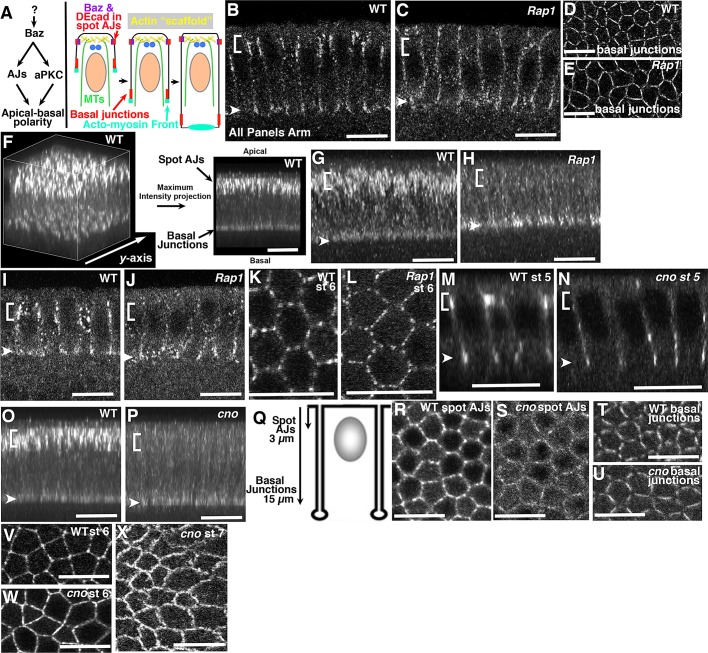
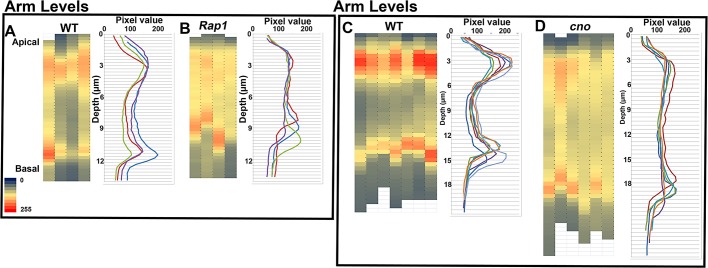
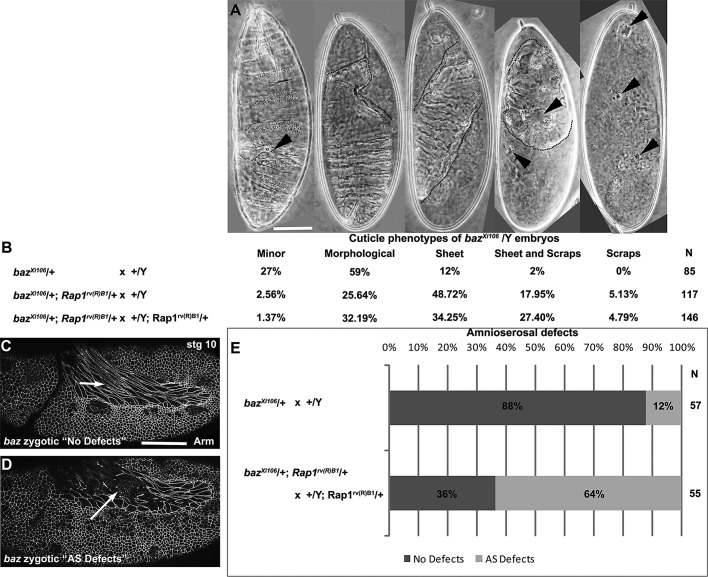
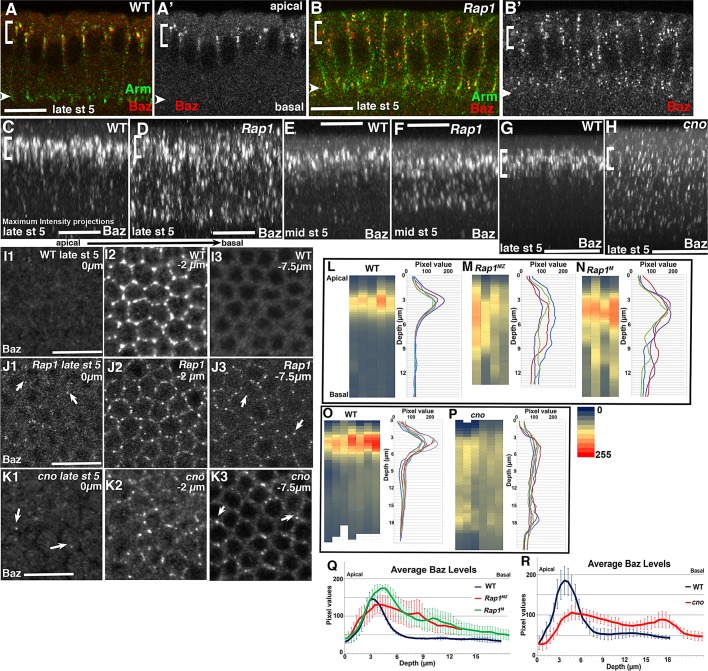
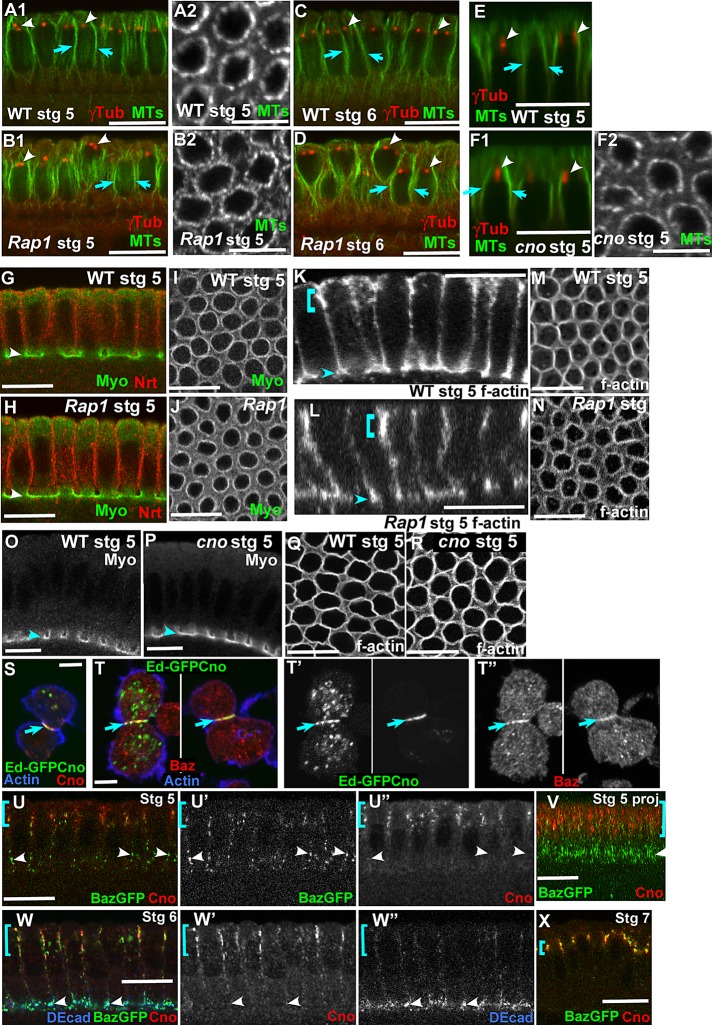
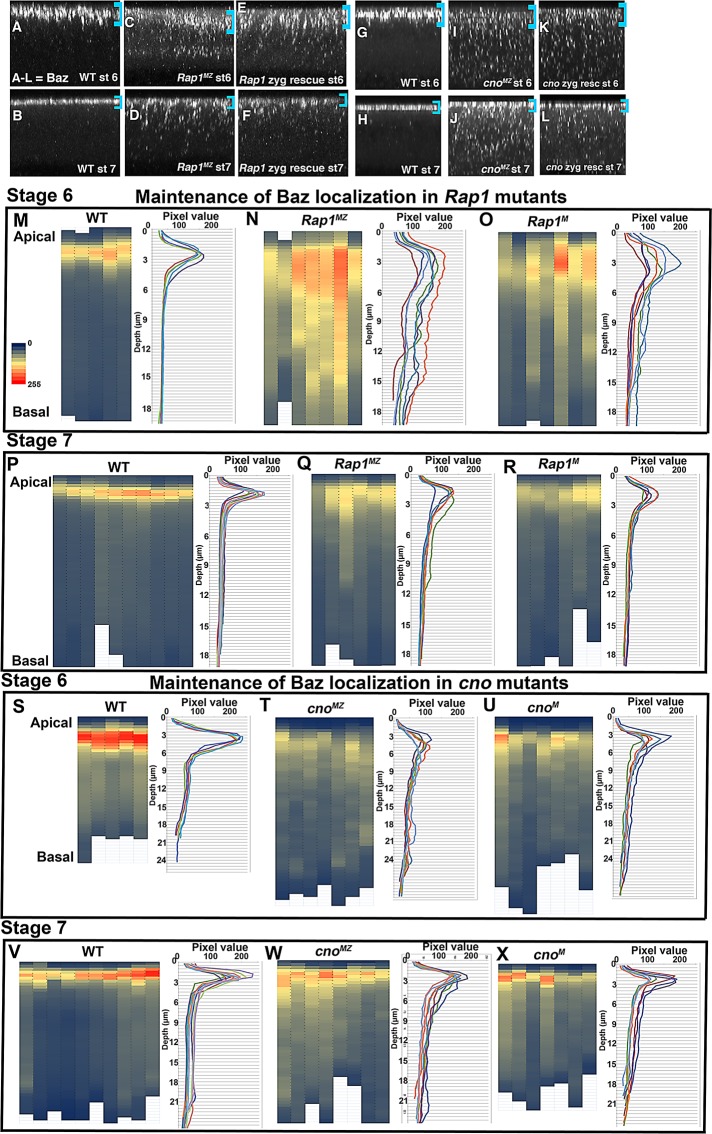
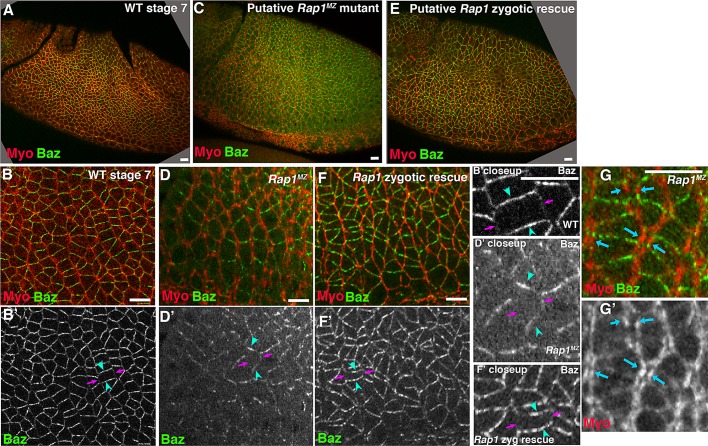
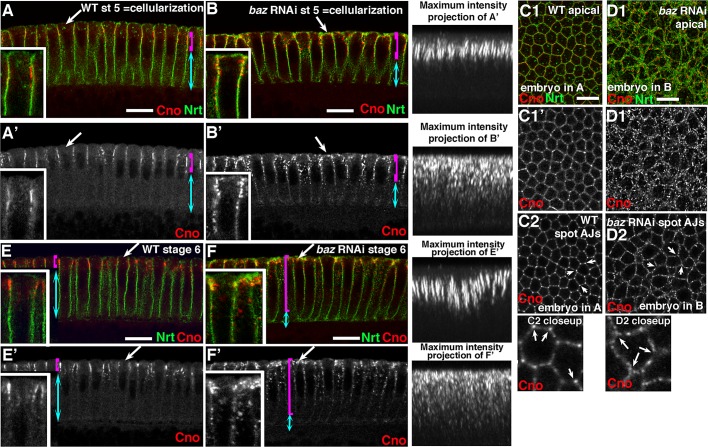
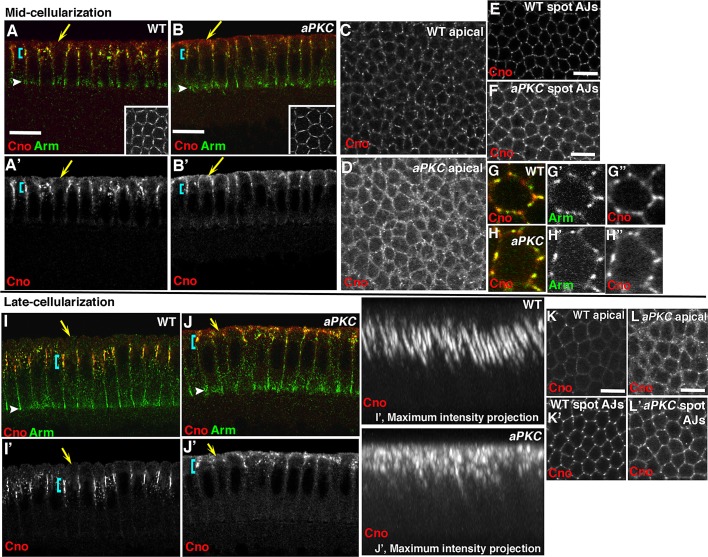
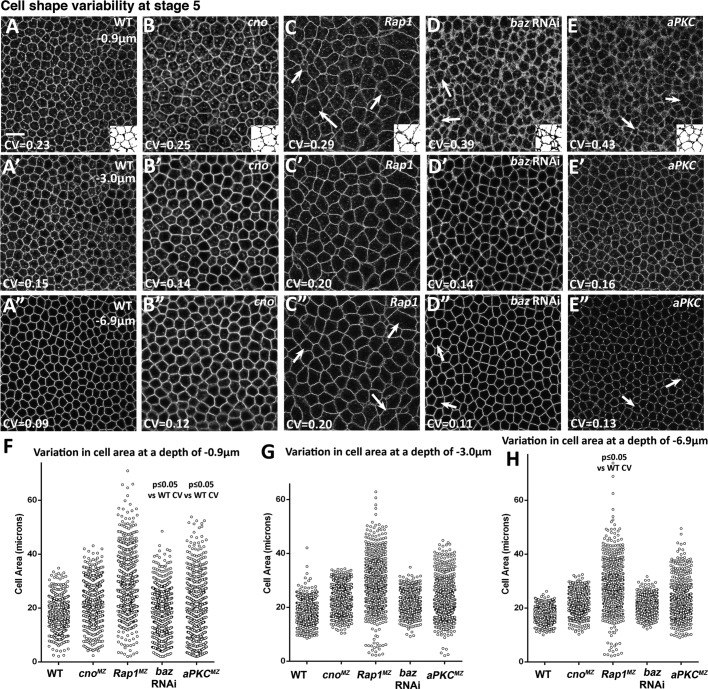
The establishment and maintenance of apical-basal cell polarity is critical for assembling epithelia and maintaining organ architecture. Drosophila embryos provide a superb model. In the current view, apically positioned Bazooka/Par3 is the initial polarity cue as cells form during cellularization. Bazooka then helps to position both adherens junctions and atypical protein kinase C (aPKC). Although a polarized cytoskeleton is critical for Bazooka positioning, proteins mediating this remained unknown. We found that the small GTPase Rap1 and the actin-junctional linker Canoe/afadin are essential for polarity establishment, as both adherens junctions and Bazooka are mispositioned in their absence. Rap1 and Canoe do not simply organize the cytoskeleton, as actin and microtubules become properly polarized in their absence. Canoe can recruit Bazooka when ectopically expressed, but they do not obligatorily colocalize. Rap1 and Canoe play continuing roles in Bazooka localization during gastrulation, but other polarity cues partially restore apical Bazooka in the absence of Rap1 or Canoe. We next tested the current linear model for polarity establishment. Both Bazooka and aPKC regulate Canoe localization despite being "downstream" of Canoe. Further, Rap1, Bazooka, and aPKC, but not Canoe, regulate columnar cell shape. These data reshape our view, suggesting that polarity establishment is regulated by a protein network rather than a linear pathway.
Figures
Similar articles
-
Rap1 acts via multiple mechanisms to position Canoe and adherens junctions and mediate apical-basal polarity establishment.Development. 2018 Jan 26;145(2):dev157941. doi: 10.1242/dev.157941. Development. 2018. PMID: 29361565 Free PMC article.
-
A contractile actomyosin network linked to adherens junctions by Canoe/afadin helps drive convergent extension.Mol Biol Cell. 2011 Jul 15;22(14):2491-508. doi: 10.1091/mbc.E11-05-0411. Epub 2011 May 25. Mol Biol Cell. 2011. PMID: 21613546 Free PMC article.
-
Rap1, Canoe and Mbt cooperate with Bazooka to promote zonula adherens assembly in the fly photoreceptor.J Cell Sci. 2018 Mar 26;131(6):jcs207779. doi: 10.1242/jcs.207779. J Cell Sci. 2018. PMID: 29507112 Free PMC article.
-
Par-3 family proteins in cell polarity & adhesion.FEBS J. 2022 Feb;289(3):596-613. doi: 10.1111/febs.15754. Epub 2021 Mar 3. FEBS J. 2022. PMID: 33565714 Free PMC article. Review.
-
The Par3/Par6/aPKC complex and epithelial cell polarity.Exp Cell Res. 2013 Jun 10;319(10):1357-64. doi: 10.1016/j.yexcr.2013.03.021. Epub 2013 Mar 25. Exp Cell Res. 2013. PMID: 23535009 Review.
Cited by
-
The Drosophila Afadin and ZO-1 homologues Canoe and Polychaetoid act in parallel to maintain epithelial integrity when challenged by adherens junction remodeling.Mol Biol Cell. 2019 Jul 22;30(16):1938-1960. doi: 10.1091/mbc.E19-04-0209. Epub 2019 Jun 12. Mol Biol Cell. 2019. PMID: 31188739 Free PMC article.
-
Redundant regulation of localization and protein stability of DmPar3.Cell Mol Life Sci. 2018 Sep;75(17):3269-3282. doi: 10.1007/s00018-018-2792-1. Epub 2018 Mar 10. Cell Mol Life Sci. 2018. PMID: 29523893 Free PMC article.
-
EGFR-dependent actomyosin patterning coordinates morphogenetic movements between tissues.bioRxiv [Preprint]. 2023 Dec 22:2023.12.22.573057. doi: 10.1101/2023.12.22.573057. bioRxiv. 2023. Update in: Dev Cell. 2025 Jan 20;60(2):270-287.e6. doi: 10.1016/j.devcel.2024.10.002. PMID: 38187543 Free PMC article. Updated. Preprint.
-
Rap1 acts via multiple mechanisms to position Canoe and adherens junctions and mediate apical-basal polarity establishment.Development. 2018 Jan 26;145(2):dev157941. doi: 10.1242/dev.157941. Development. 2018. PMID: 29361565 Free PMC article.
-
EGFR-dependent actomyosin patterning coordinates morphogenetic movements between tissues in Drosophila melanogaster.Dev Cell. 2025 Jan 20;60(2):270-287.e6. doi: 10.1016/j.devcel.2024.10.002. Epub 2024 Oct 25. Dev Cell. 2025. PMID: 39461341
References
-
- Assemat E, Bazellieres E, Pallesi-Pocachard E, Le Bivic A, Massey-Harroche D. Polarity complex proteins. Biochim Biophys Acta. 2008;1778:614–630. - PubMed
-
- Bertet C, Sulak L, Lecuit T. Myosin-dependent junction remodelling controls planar cell intercalation and axis elongation. Nature. 2004;429:667–671. - PubMed
-
- Bilder D, Schober M, Perrimon N. Integrated activity of PDZ protein complexes regulates epithelial polarity. Nat Cell Biol. 2003;5:53–58. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
