

SNF5 reexpression in malignant rhabdoid tumors regulates transcription of target genes by recruitment of SWI/SNF complexes and RNAPII to the transcription start site of their promoters
- PMID: 23364536
- PMCID: PMC4342046
- DOI: 10.1158/1541-7786.MCR-12-0390
SNF5 reexpression in malignant rhabdoid tumors regulates transcription of target genes by recruitment of SWI/SNF complexes and RNAPII to the transcription start site of their promoters
Abstract
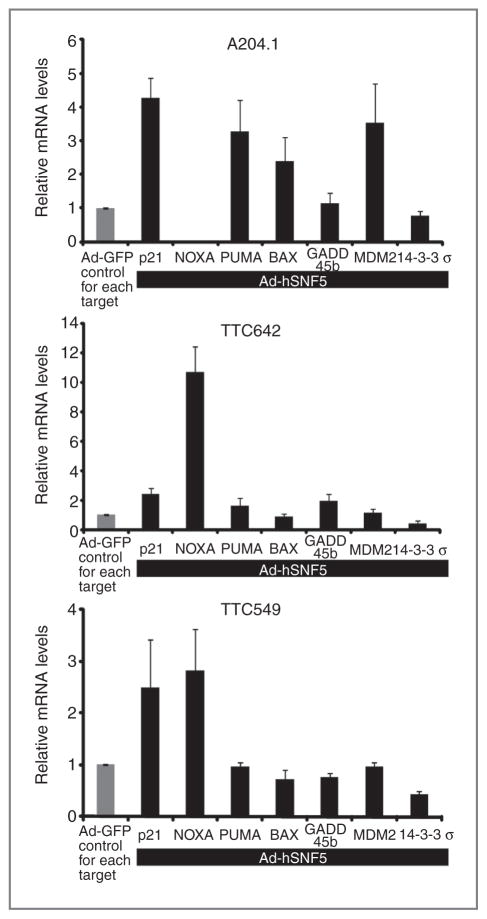
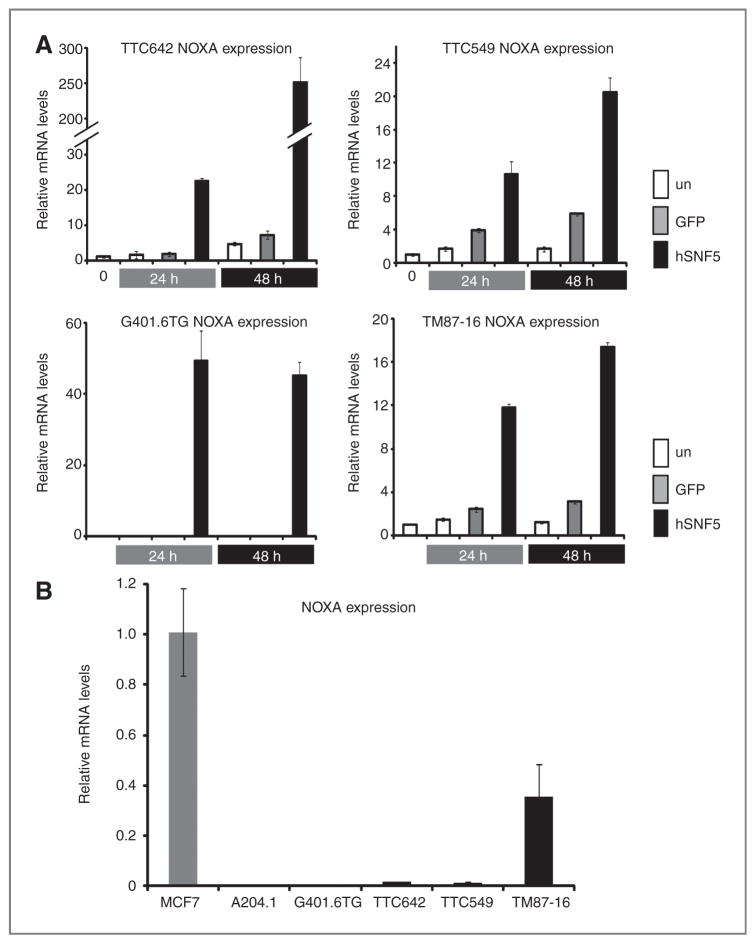
Malignant rhabdoid tumor (MRT), a highly aggressive cancer of young children, displays inactivation or loss of the hSNF5/INI1/SMARCB1 gene, a core subunit of the SWI/SNF chromatin-remodeling complex, in primary tumors and cell lines. We have previously reported that reexpression of hSNF5 in some MRT cell lines causes a G1 arrest via p21(CIP1/WAF1) (p21) mRNA induction in a p53-independent manner. However, the mechanism(s) by which hSNF5 reexpression activates gene transcription remains unclear. We initially searched for other hSNF5 target genes by asking whether hSNF5 loss altered regulation of other consensus p53 target genes. Our studies show that hSNF5 regulates only a subset of p53 target genes, including p21 and NOXA, in MRT cell lines. We also show that hSNF5 reexpression modulates SWI/SNF complex levels at the transcription start site (TSS) at both loci and leads to activation of transcription initiation through recruitment of RNA polymerase II (RNAPII) accompanied by H3K4 and H3K36 modifications. Furthermore, our results show lower NOXA expression in MRT cell lines compared with other human tumor cell lines, suggesting that hSNF5 loss may alter the expression of this important apoptotic gene. Thus, one mechanism for MRT development after hSNF5 loss may rely on reduced chromatin-remodeling activity of the SWI/SNF complex at the TSS of critical gene promoters. Furthermore, because we observe growth inhibition after NOXA expression in MRT cells, the NOXA pathway may provide a novel target with clinical relevancy for treatment of this aggressive disease.
Conflict of interest statement
No potential conflicts of interest were disclosed.
Figures



References
-
- Beckwith JB, Palmer NF. Histopathology and prognosis of Wilms tumors: results from the First National Wilms’ Tumor Study. Cancer. 1978;41:1937–48. - PubMed
-
- Biegel JA, Tan L, Zhang F, Wainwright L, Russo P, Rorke LB. Alterations of the hSNF5/INI1 gene in central nervous system atypical teratoid/rhabdoid tumors and renal and extrarenal rhabdoid tumors. Clin Cancer Res. 2002;8:3461–7. - PubMed
-
- Hoot AC, Russo P, Judkins AR, Perlman EJ, Biegel JA. Immunohistochemical analysis of hSNF5/INI1 distinguishes renal and extra-renal malignant rhabdoid tumors from other pediatric soft tissue tumors. Am J Surg Pathol. 2004;28:1485–91. - PubMed
-
- Tomlinson GE, Breslow NE, Dome J, Guthrie KA, Norkool P, Li S, et al. Rhabdoid tumor of the kidney in the National Wilms’ Tumor Study: age at diagnosis as a prognostic factor. J Clin Oncol. 2005;23:7641–5. - PubMed
-
- Versteege I, Sevenet N, Lange J, Rousseau-Merck MF, Ambros P, Handgretinger R, et al. Truncating mutations of hSNF5/INI1 in aggressive paediatric cancer. Nature. 1998;394:203–6. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
