

tmc-1 encodes a sodium-sensitive channel required for salt chemosensation in C. elegans
- PMID: 23364694
- PMCID: PMC4021456
- DOI: 10.1038/nature11845
tmc-1 encodes a sodium-sensitive channel required for salt chemosensation in C. elegans
Abstract
Transmembrane channel-like (TMC) genes encode a broadly conserved family of multipass integral membrane proteins in animals. Human TMC1 and TMC2 genes are linked to human deafness and required for hair-cell mechanotransduction; however, the molecular functions of these and other TMC proteins have not been determined. Here we show that the Caenorhabditis elegans tmc-1 gene encodes a sodium sensor that functions specifically in salt taste chemosensation. tmc-1 is expressed in the ASH polymodal avoidance neurons, where it is required for salt-evoked neuronal activity and behavioural avoidance of high concentrations of NaCl. However, tmc-1 has no effect on responses to other stimuli sensed by the ASH neurons including high osmolarity and chemical repellents, indicating a specific role in salt sensation. When expressed in mammalian cell culture, C. elegans TMC-1 generates a predominantly cationic conductance activated by high extracellular sodium but not by other cations or uncharged small molecules. Thus, TMC-1 is both necessary for salt sensation in vivo and sufficient to generate a sodium-sensitive channel in vitro, identifying it as a probable ionotropic sensory receptor.
Figures
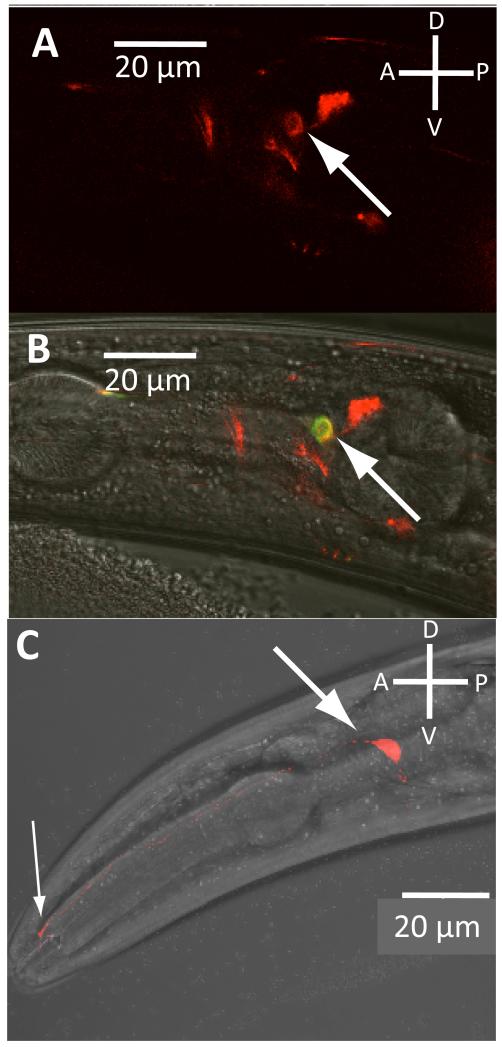
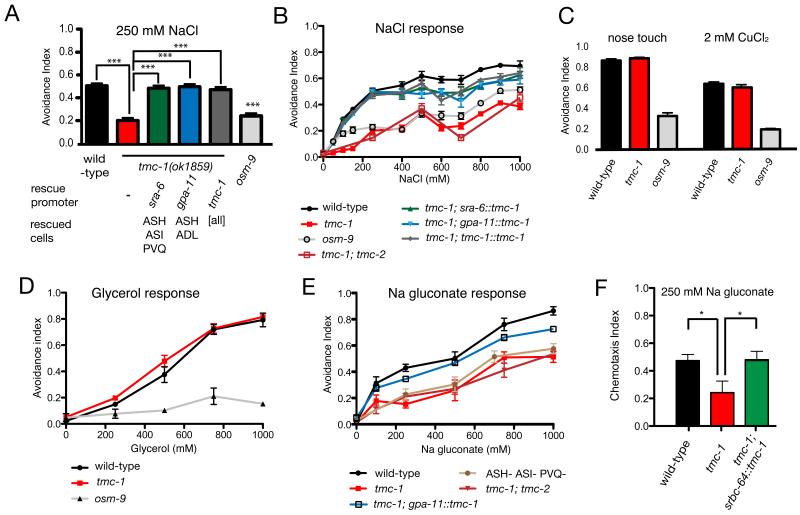
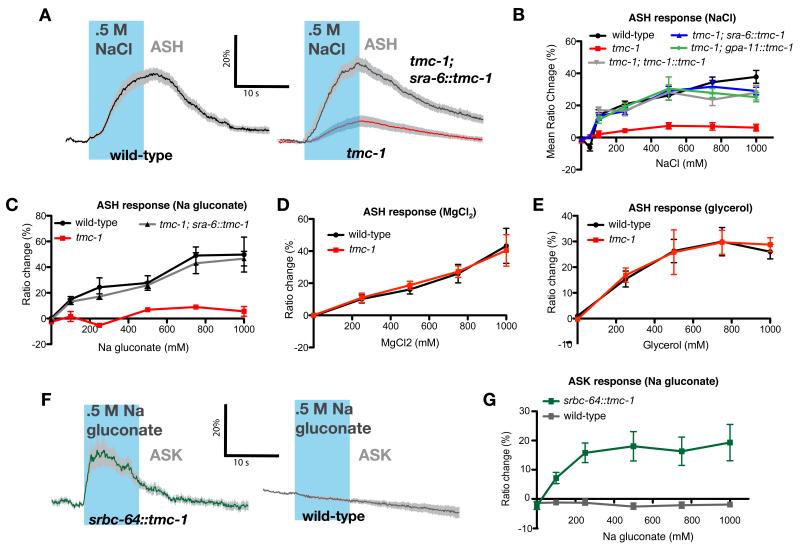
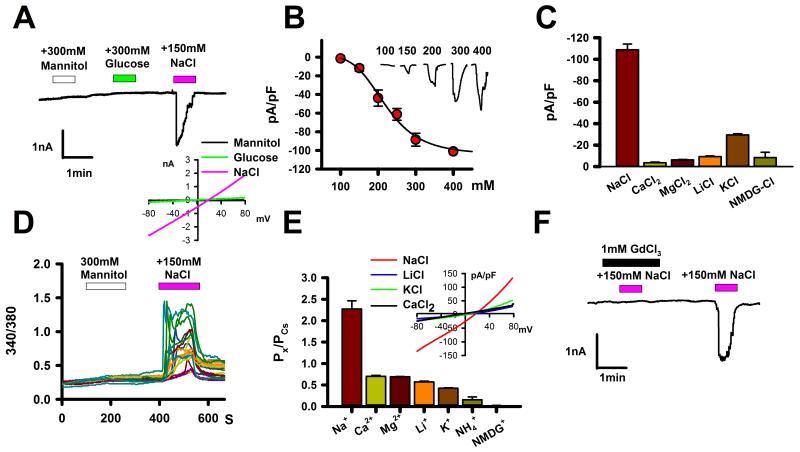
Comment in
-
Neuroscience: Salty sensations.Nature. 2013 Feb 7;494(7435):44-5. doi: 10.1038/nature11946. Epub 2013 Jan 30. Nature. 2013. PMID: 23364688 No abstract available.
References
-
- Kurima K, Yang Y, Sorber K, Griffith AJ. Characterization of the transmembrane channel-like (TMC) gene family: functional clues from hearing loss and epidermodysplasia verruciformis. Genomics. 2003;82:300–308. - PubMed
-
- Kurima K, et al. Dominant and recessive deafness caused by mutations of a novel gene, TMC1, required for cochlear hair-cell function. Nature genetics. 2002;30:277–284. doi:10.1038/ng842. - PubMed
-
- Vreugde S, et al. Beethoven, a mouse model for dominant, progressive hearing loss DFNA36. Nature genetics. 2002;30:257–258. doi:10.1038/ng848. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
