

Nuclear calcium signaling regulates nuclear export of a subset of class IIa histone deacetylases following synaptic activity
- PMID: 23364788
- PMCID: PMC3605626
- DOI: 10.1074/jbc.M112.432773
Nuclear calcium signaling regulates nuclear export of a subset of class IIa histone deacetylases following synaptic activity
Abstract
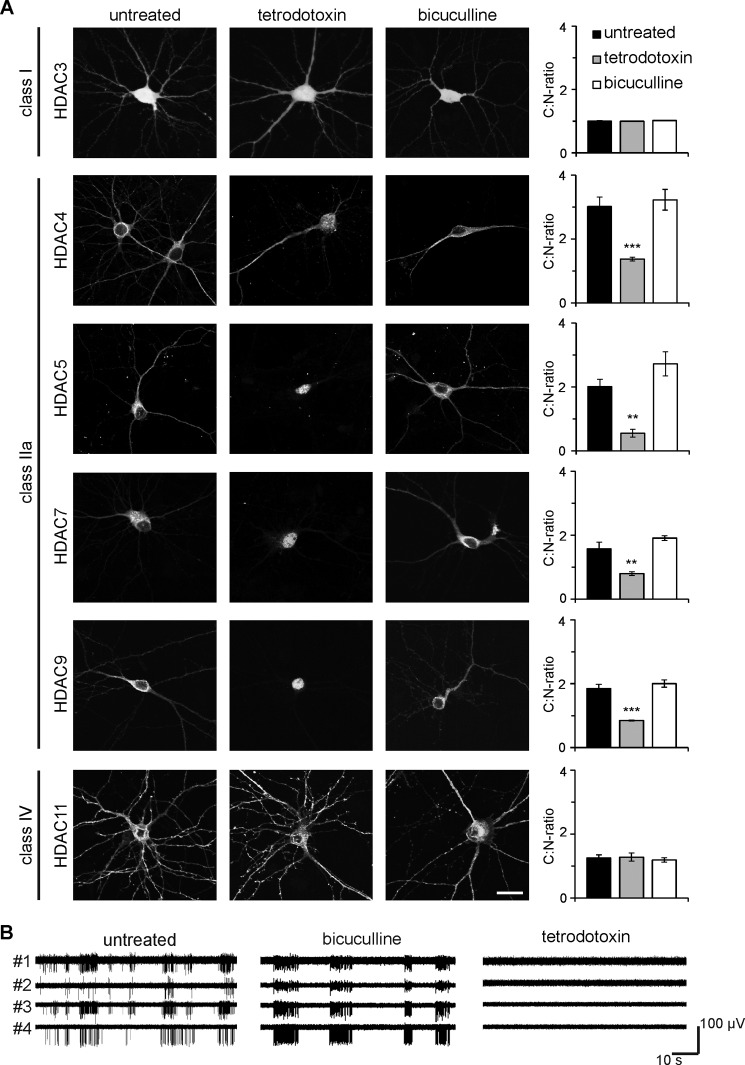
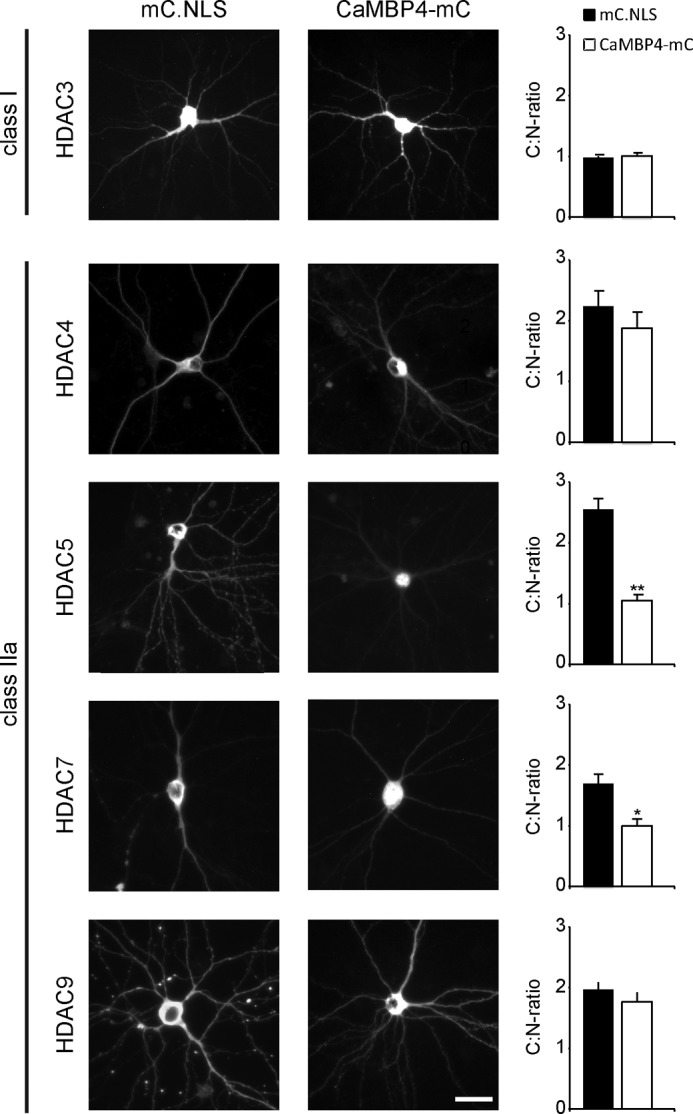
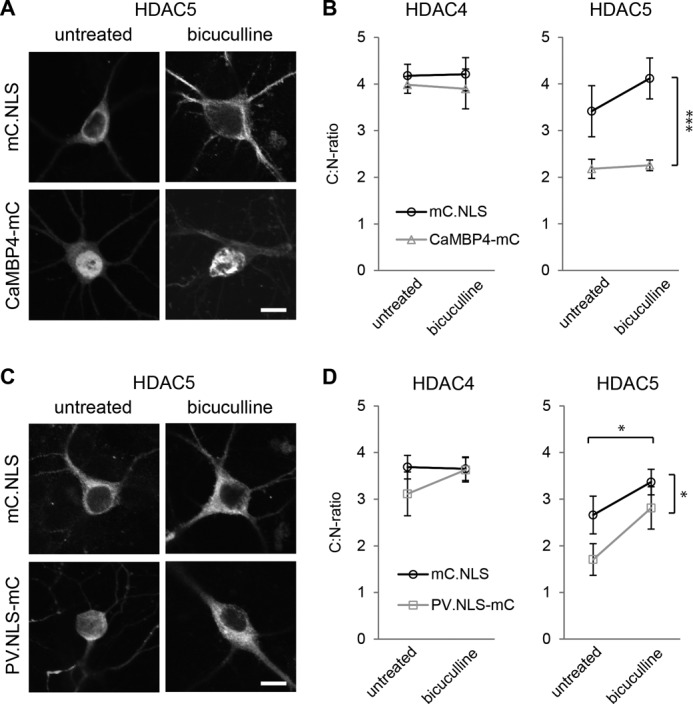
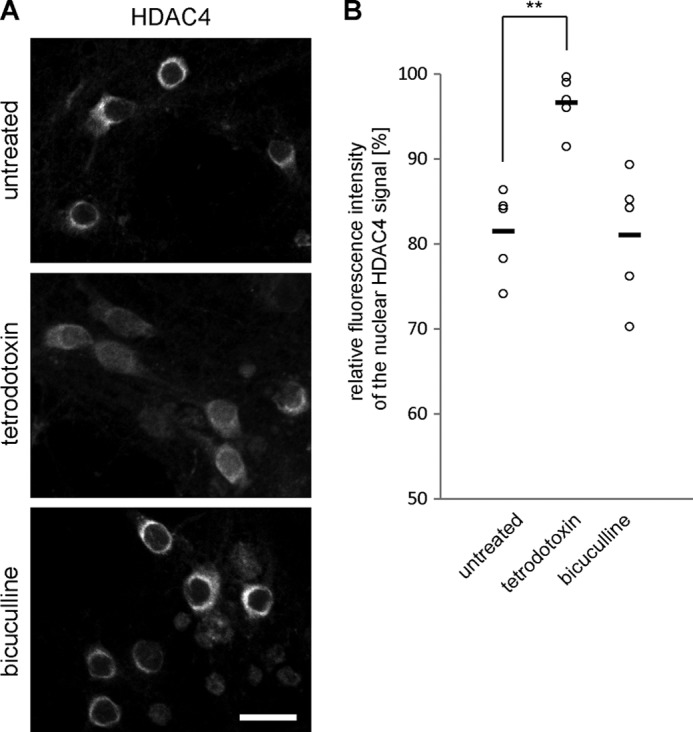
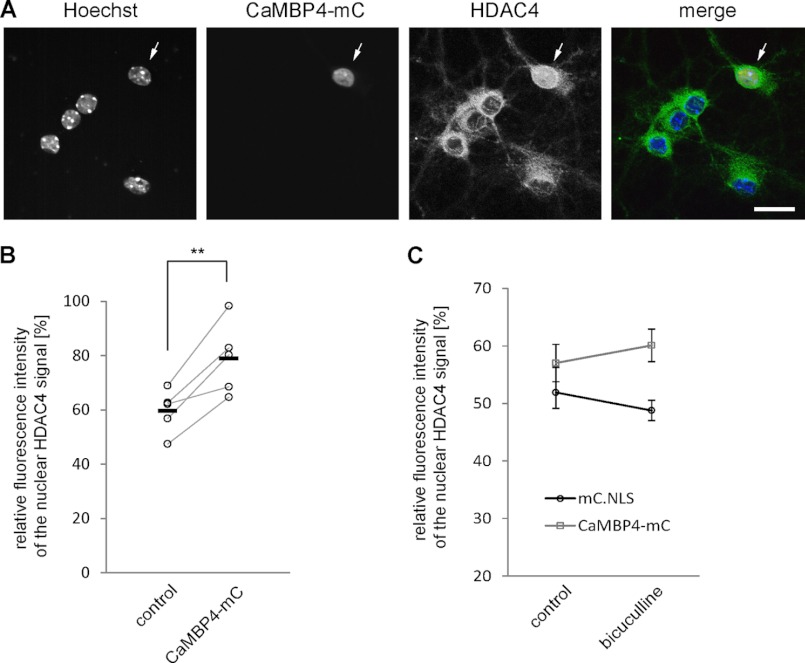
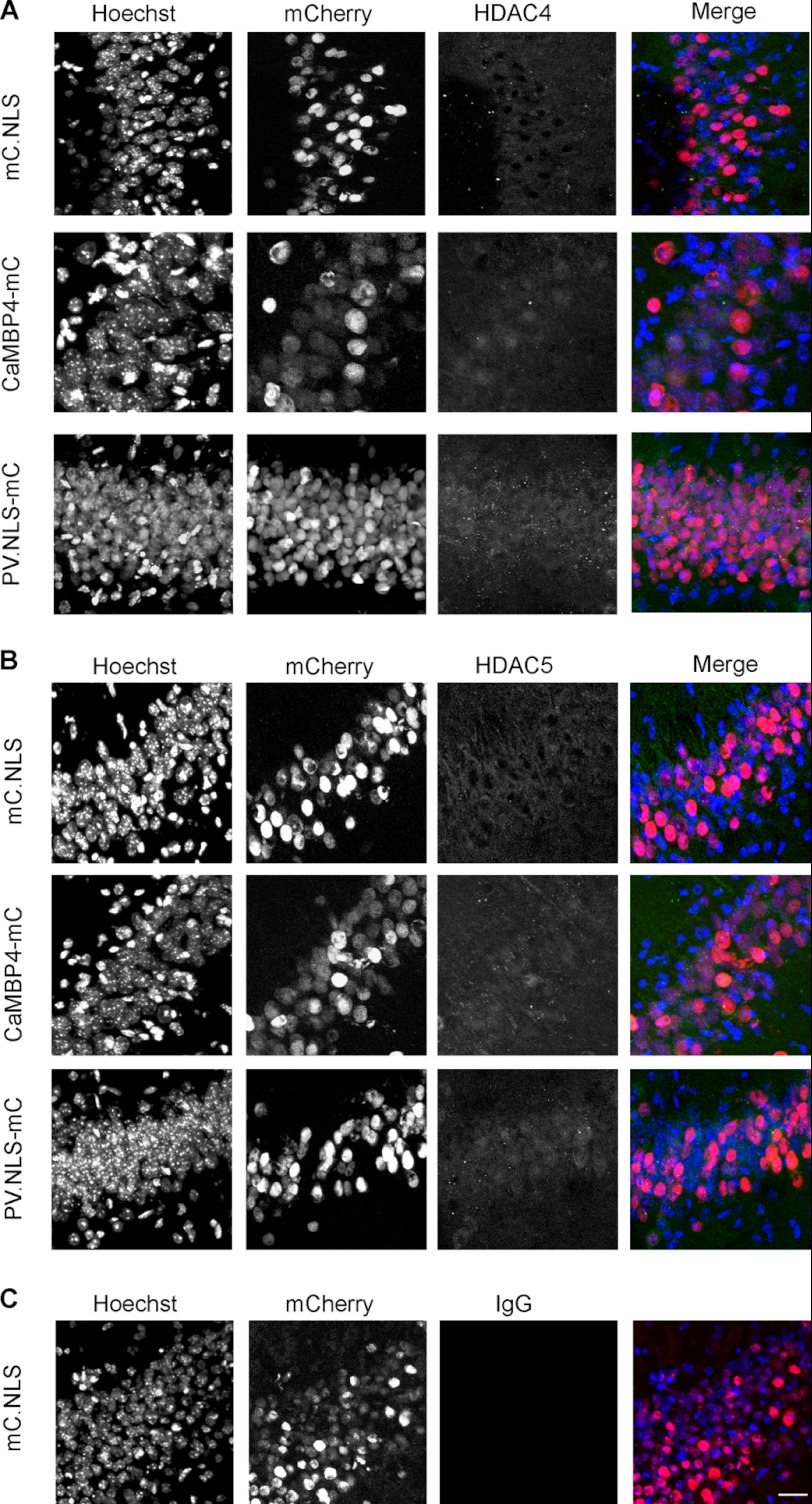
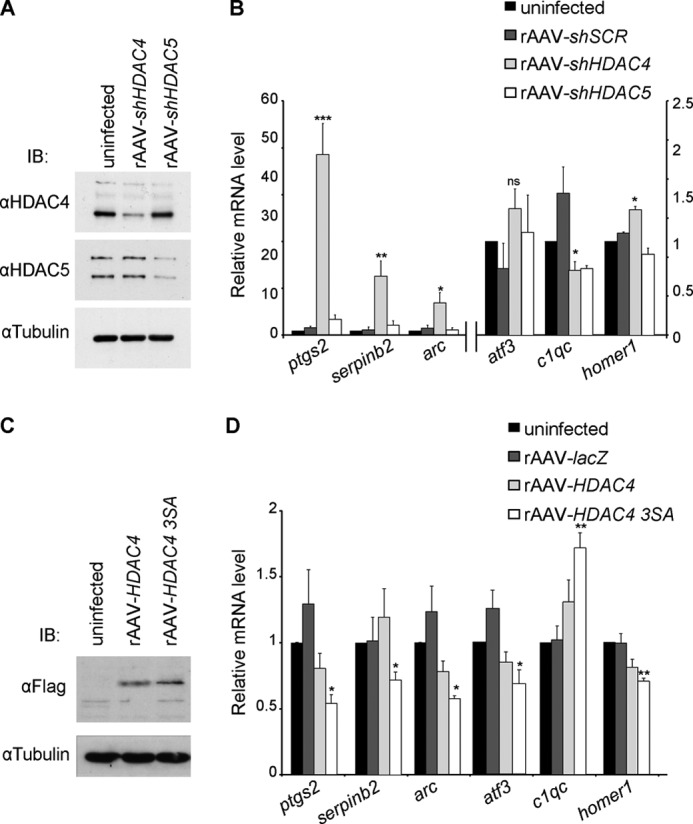
In neurons, dynamic changes in the subcellular localization of histone deacetylases (HDACs) are thought to contribute to signal-regulated gene expression. Here we show that in mouse hippocampal neurons, synaptic activity-dependent nucleo-cytoplasmic shuttling is a common feature of all members of class IIa HDACs, which distinguishes them from other classes of HDACs. Nuclear calcium, a key regulator in neuronal gene expression, is required for the nuclear export of a subset of class IIa HDACs. We found that inhibition of nuclear calcium signaling using CaMBP4 or increasing the nuclear calcium buffering capacity by means of expression of a nuclear targeted version of parvalbumin (PV.NLS-mC) led to a build-up of HDAC4 and HDAC5 in the cell nucleus, which in the case of PV.NLS-mC can be reversed by nuclear calcium transients triggered by bursts of action potential firing. A similar nuclear accumulation of HDAC4 and HDAC5 was observed in vivo in the mouse hippocampus following stereotaxic delivery of recombinant adeno-associated viruses expressing either CaMBP4 or PV.NLS-mC. The modulation of HDAC4 activity either by RNA interference-mediated reduction of HDAC4 protein levels or by expression of a constitutively nuclear localized mutant of HDAC4 leads to changes in the mRNA levels of several nuclear calcium-regulated genes with known functions in acquired neuroprotection (atf3, serpinb2), memory consolidation (homer1, arc), and the development of chronic pain (ptgs2, c1qc). These results identify nuclear calcium as a regulator of nuclear export of HDAC4 and HDAC5. The reduction of nuclear localized HDACs represents a novel transcription-promoting pathway stimulated by nuclear calcium.
Figures
Similar articles
-
Neuronal activity-dependent nucleocytoplasmic shuttling of HDAC4 and HDAC5.J Neurochem. 2003 Apr;85(1):151-9. doi: 10.1046/j.1471-4159.2003.01648.x. J Neurochem. 2003. PMID: 12641737
-
Histone deacetylase 4 shapes neuronal morphology via a mechanism involving regulation of expression of vascular endothelial growth factor D.J Biol Chem. 2018 May 25;293(21):8196-8207. doi: 10.1074/jbc.RA117.001613. Epub 2018 Apr 9. J Biol Chem. 2018. PMID: 29632070 Free PMC article.
-
14-3-3 regulates the nuclear import of class IIa histone deacetylases.Biochem Biophys Res Commun. 2008 Dec 19;377(3):852-6. doi: 10.1016/j.bbrc.2008.10.079. Epub 2008 Oct 24. Biochem Biophys Res Commun. 2008. PMID: 18952052
-
The Class IIa histone deacetylase HDAC4 and neuronal function: Nuclear nuisance and cytoplasmic stalwart?Neurobiol Learn Mem. 2015 Sep;123:149-58. doi: 10.1016/j.nlm.2015.06.006. Epub 2015 Jun 11. Neurobiol Learn Mem. 2015. PMID: 26074448 Review.
-
Class IIa HDACs - new insights into their functions in physiology and pathology.FEBS J. 2015 May;282(9):1736-44. doi: 10.1111/febs.13061. Epub 2014 Oct 27. FEBS J. 2015. PMID: 25244360 Review.
Cited by
-
The nAChR Chaperone TMEM35a (NACHO) Contributes to the Development of Hyperalgesia in Mice.Neuroscience. 2021 Mar 1;457:74-87. doi: 10.1016/j.neuroscience.2020.12.027. Epub 2021 Jan 7. Neuroscience. 2021. PMID: 33422618 Free PMC article.
-
VEGFD signaling balances stability and activity-dependent structural plasticity of dendrites.Cell Mol Life Sci. 2024 Aug 19;81(1):354. doi: 10.1007/s00018-024-05357-2. Cell Mol Life Sci. 2024. PMID: 39158743 Free PMC article.
-
Histone deacetylase expression patterns in developing murine optic nerve.BMC Dev Biol. 2014 Jul 9;14:30. doi: 10.1186/1471-213X-14-30. BMC Dev Biol. 2014. PMID: 25011550 Free PMC article.
-
Histone Deacetylases Contribute to Excitotoxicity-Triggered Degeneration of Retinal Ganglion Cells In Vivo.Mol Neurobiol. 2019 Dec;56(12):8018-8034. doi: 10.1007/s12035-019-01658-x. Epub 2019 Jun 3. Mol Neurobiol. 2019. PMID: 31161423
-
Nesprin-2 mediated nuclear trafficking and its clinical implications.Nucleus. 2015;6(6):479-89. doi: 10.1080/19491034.2015.1128608. Epub 2015 Dec 8. Nucleus. 2015. PMID: 26645154 Free PMC article.
References
-
- Greer P. L., Greenberg M. E. (2008) From synapse to nucleus: calcium-dependent gene transcription in the control of synapse development and function. Neuron 59, 846–860 - PubMed
-
- Hardingham G. E., Chawla S., Johnson C. M., Bading H. (1997) Distinct functions of nuclear and cytoplasmic calcium in the control of gene expression. Nature 385, 260–265 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
