

Epstein-Barr virus deubiquitinase downregulates TRAF6-mediated NF-κB signaling during productive replication
- PMID: 23365429
- PMCID: PMC3624234
- DOI: 10.1128/JVI.02020-12
Epstein-Barr virus deubiquitinase downregulates TRAF6-mediated NF-κB signaling during productive replication
Abstract
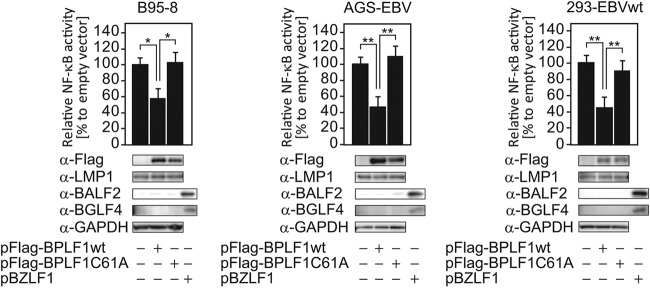
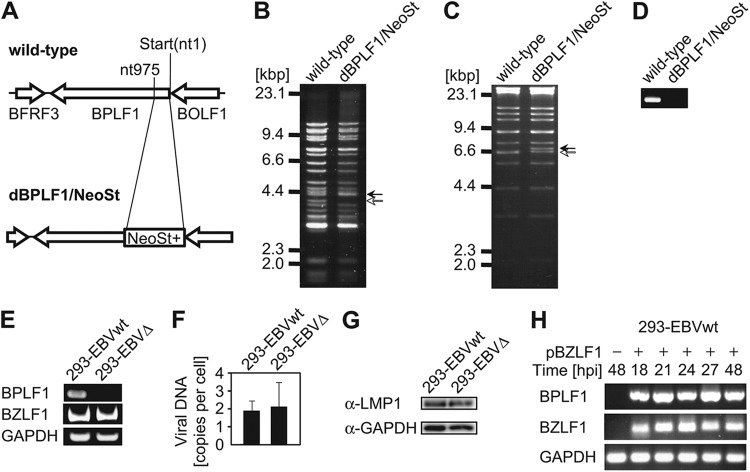
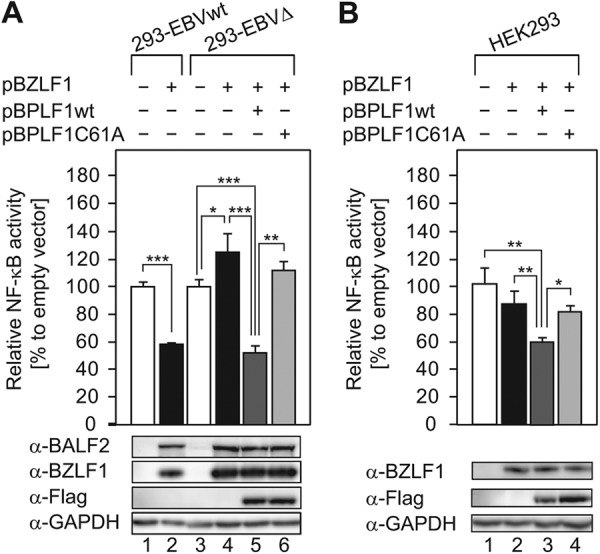
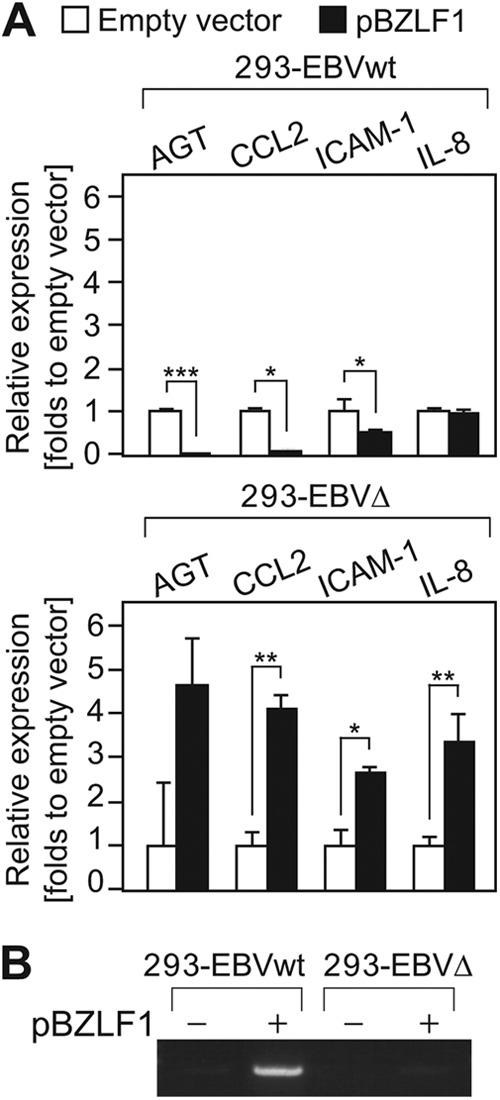
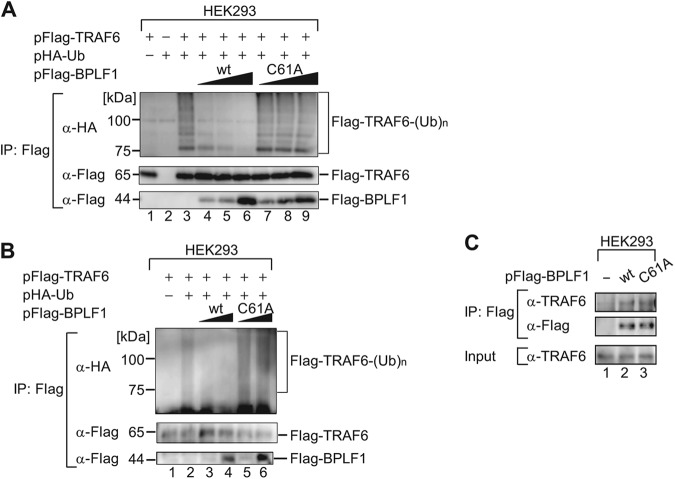
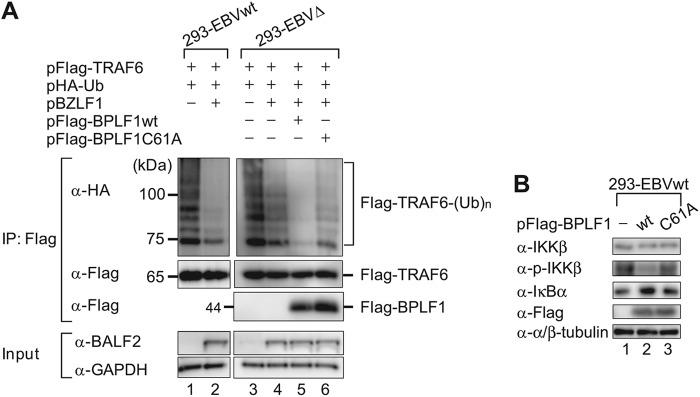
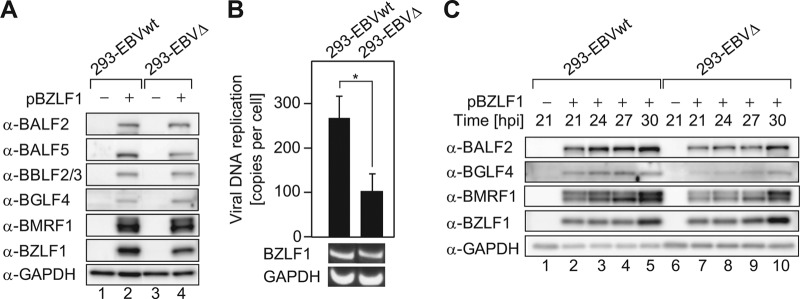
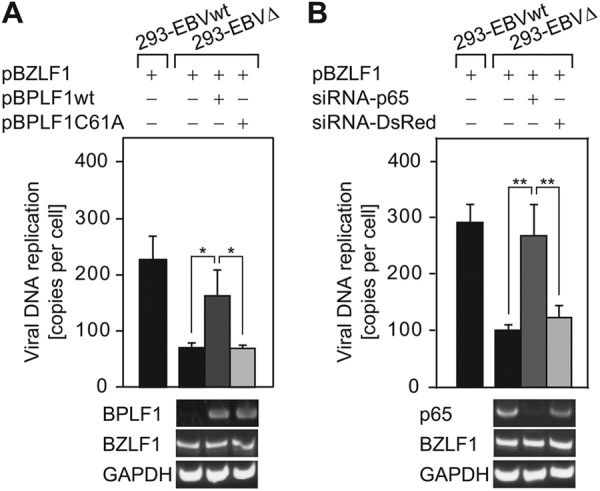
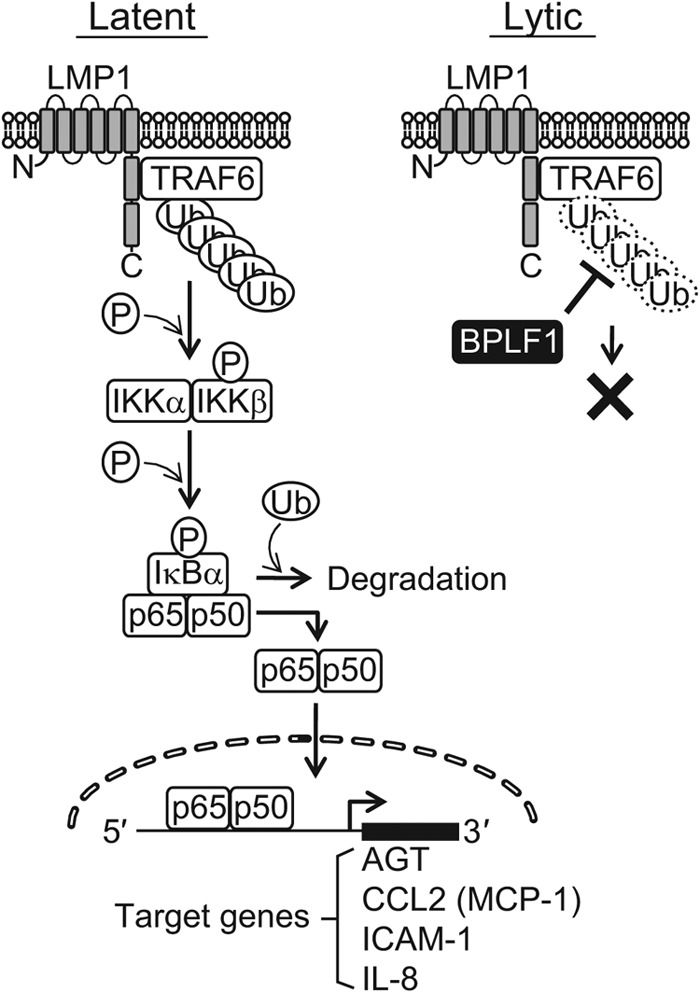
Epstein-Barr virus (EBV), a human oncogenic herpesvirus that establishes a lifelong latent infection in the host, occasionally enters lytic infection to produce progeny viruses. The EBV oncogene latent membrane protein 1 (LMP1), which is expressed in both latent and lytic infection, constitutively activates the canonical NF-κB (p65) pathway. Such LMP1-mediated NF-κB activation is necessary for proliferation of latently infected cells and inhibition of viral lytic cycle progression. Actually, canonical NF-κB target gene expression was suppressed upon the onset of lytic infection. TRAF6, which is activated by conjugation of polyubiquitin chains, associates with LMP1 to mediate NF-κB signal transduction. We have found that EBV-encoded BPLF1 interacts with and deubiquitinates TRAF6 to inhibit NF-κB signaling during lytic infection. HEK293 cells with BPLF1-deficient recombinant EBV exhibited poor viral DNA replication compared with the wild type. Furthermore, exogenous expression of BPLF1 or p65 knockdown in cells restored DNA replication of BPLF1-deficient viruses, indicating that EBV BPLF1 deubiquitinates TRAF6 to inhibit NF-κB signal transduction, leading to promotion of viral lytic DNA replication.
Figures
Similar articles
-
The Rad6/18 ubiquitin complex interacts with the Epstein-Barr virus deubiquitinating enzyme, BPLF1, and contributes to virus infectivity.J Virol. 2014 Jun;88(11):6411-22. doi: 10.1128/JVI.00536-14. Epub 2014 Mar 26. J Virol. 2014. PMID: 24672041 Free PMC article.
-
Epstein-Barr virus large tegument protein BPLF1 contributes to innate immune evasion through interference with toll-like receptor signaling.PLoS Pathog. 2014 Feb 20;10(2):e1003960. doi: 10.1371/journal.ppat.1003960. eCollection 2014 Feb. PLoS Pathog. 2014. PMID: 24586164 Free PMC article.
-
The C-terminal activating region 2 of the Epstein-Barr virus-encoded latent membrane protein 1 activates NF-kappaB through TRAF6 and TAK1.J Biol Chem. 2006 Jan 27;281(4):2162-9. doi: 10.1074/jbc.M505903200. Epub 2005 Nov 8. J Biol Chem. 2006. PMID: 16280329
-
NF-κB and IRF7 pathway activation by Epstein-Barr virus Latent Membrane Protein 1.Viruses. 2013 Jun 21;5(6):1587-606. doi: 10.3390/v5061587. Viruses. 2013. PMID: 23793113 Free PMC article. Review.
-
LMP1 TRAFficking activates growth and survival pathways.Adv Exp Med Biol. 2007;597:173-87. doi: 10.1007/978-0-387-70630-6_14. Adv Exp Med Biol. 2007. PMID: 17633026 Review.
Cited by
-
Epstein-Barr virus genetic variation in lymphoblastoid cell lines derived from Kenyan pediatric population.PLoS One. 2015 May 1;10(5):e0125420. doi: 10.1371/journal.pone.0125420. eCollection 2015. PLoS One. 2015. PMID: 25933165 Free PMC article.
-
Strategies of Epstein-Barr virus to evade innate antiviral immunity of its human host.Front Microbiol. 2022 Jul 22;13:955603. doi: 10.3389/fmicb.2022.955603. eCollection 2022. Front Microbiol. 2022. PMID: 35935191 Free PMC article. Review.
-
Essential role of HCMV deubiquitinase in promoting oncogenesis by targeting anti-viral innate immune signaling pathways.Cell Death Dis. 2017 Oct 5;8(10):e3078. doi: 10.1038/cddis.2017.461. Cell Death Dis. 2017. PMID: 28981114 Free PMC article.
-
Identifying the key regulators orchestrating Epstein-Barr virus reactivation.Front Microbiol. 2024 Dec 5;15:1505191. doi: 10.3389/fmicb.2024.1505191. eCollection 2024. Front Microbiol. 2024. PMID: 39703703 Free PMC article. Review.
-
Microbial strategies for antagonizing Toll-like-receptor signal transduction.Curr Opin Immunol. 2015 Feb;32:61-70. doi: 10.1016/j.coi.2014.12.011. Epub 2015 Jan 20. Curr Opin Immunol. 2015. PMID: 25615700 Free PMC article. Review.
References
-
- Baer R, Bankier AT, Biggin MD, Deininger PL, Farrell PJ, Gibson TJ, Hatfull G, Hudson GS, Satchwell SC, Séguin C, Tuffnell PS, Barrell BG. 1984. DNA sequence and expression of the B95-8 Epstein-Barr virus genome. Nature 310:207–211 - PubMed
-
- Morrison JA, Gulley ML, Pathmanathan R, Raab-Traub N. 2004. Differential signaling pathways are activated in the Epstein-Barr virus-associated malignancies nasopharyngeal carcinoma and Hodgkin lymphoma. Cancer Res. 64:5251–5260 - PubMed
-
- Wu S, Xie P, Welsh K, Li C, Ni CZ, Zhu X, Reed JC, Satterthwait AC, Bishop GA, Ely KR. 2005. LMP1 protein from the Epstein-Barr virus is a structural CD40 decoy in B lymphocytes for binding to TRAF3. J. Biol. Chem. 280:33620–33626 - PubMed
-
- Xie P, Bishop GA. 2004. Roles of TNF receptor-associated factor 3 in signaling to B lymphocytes by carboxyl-terminal activating regions 1 and 2 of the EBV-encoded oncoprotein latent membrane protein 1. J. Immunol. 173:5546–5555 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
