

Multi-channel acoustic recording and automated analysis of Drosophila courtship songs
- PMID: 23369160
- PMCID: PMC3599446
- DOI: 10.1186/1741-7007-11-11
Multi-channel acoustic recording and automated analysis of Drosophila courtship songs
Abstract
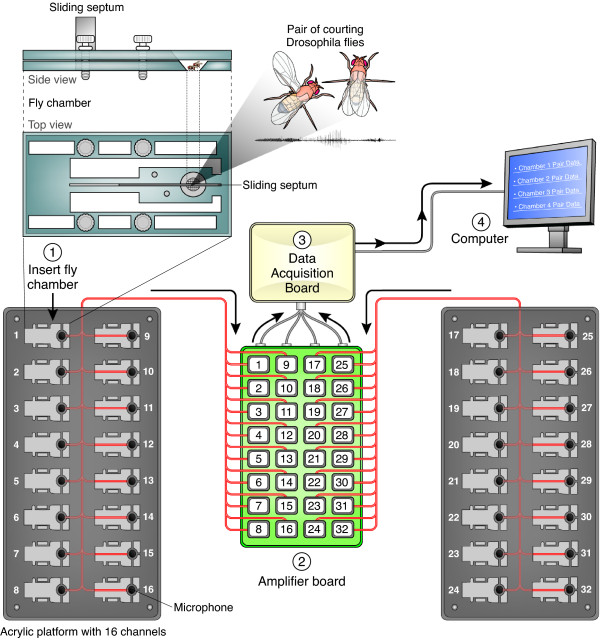
Background: Drosophila melanogaster has served as a powerful model system for genetic studies of courtship songs. To accelerate research on the genetic and neural mechanisms underlying courtship song, we have developed a sensitive recording system to simultaneously capture the acoustic signals from 32 separate pairs of courting flies as well as software for automated segmentation of songs.
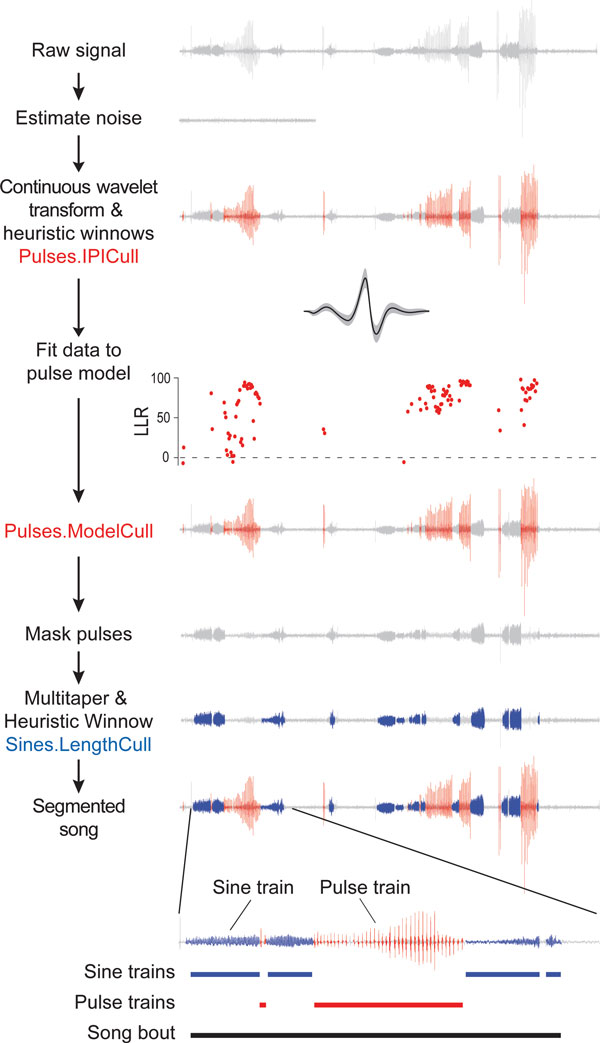
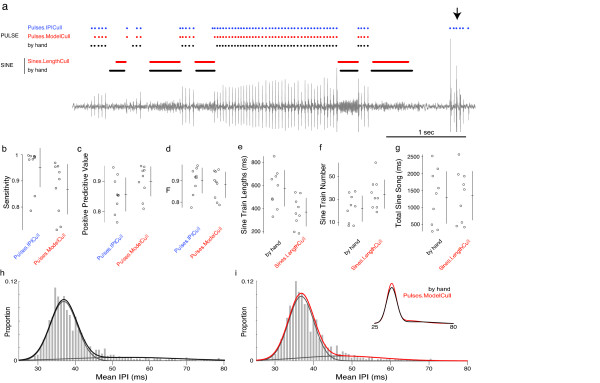
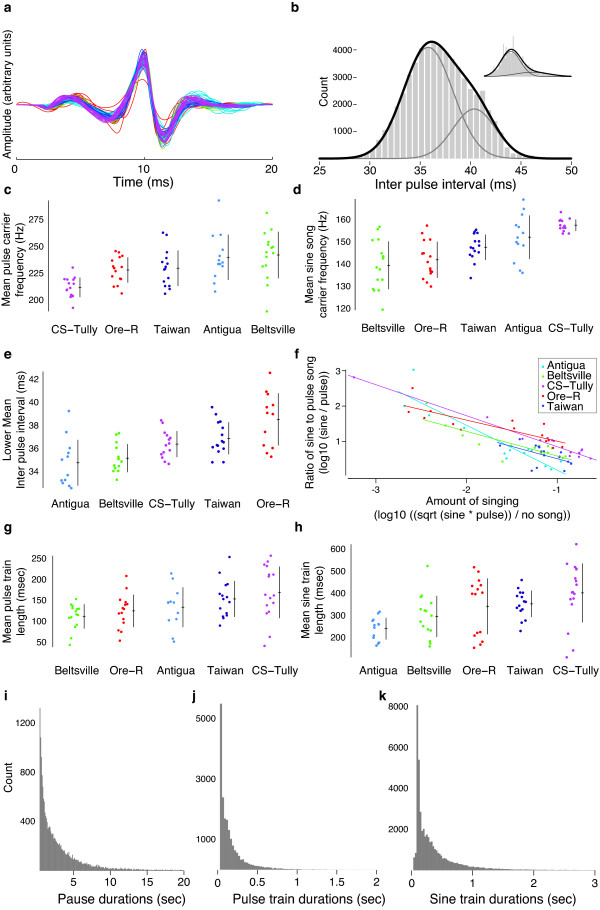
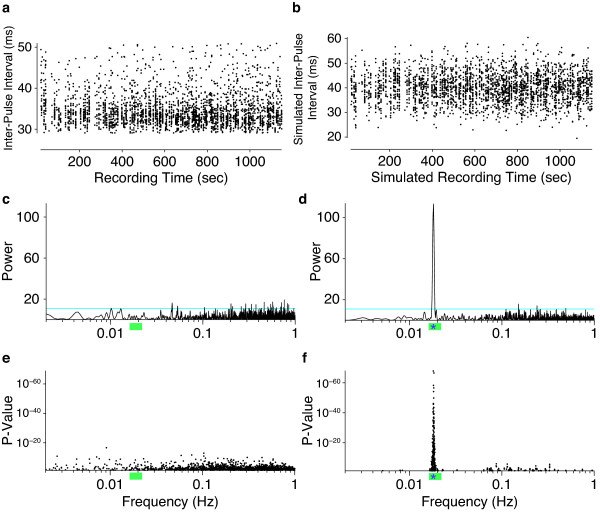
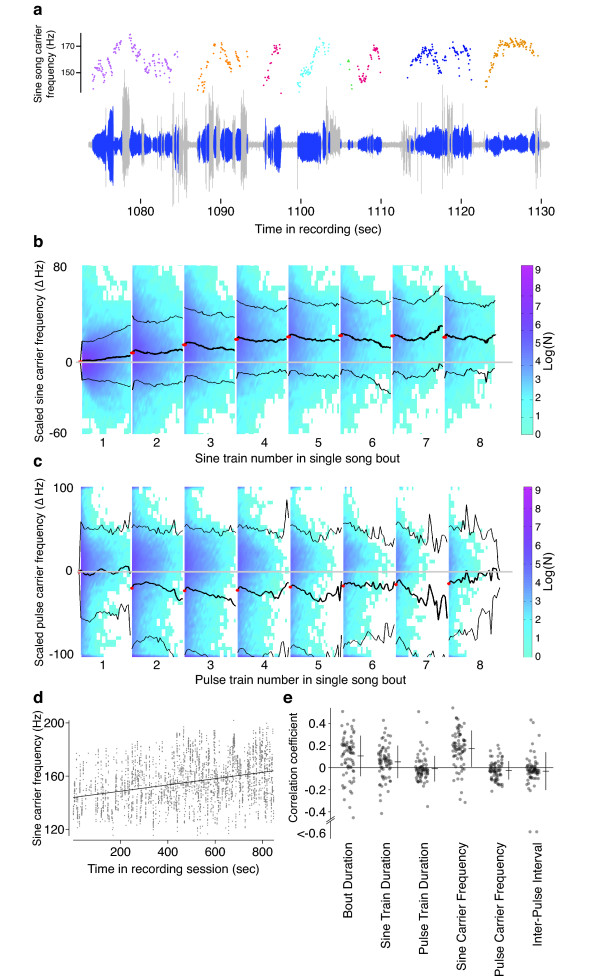
Results: Our novel hardware design enables recording of low amplitude sounds in most laboratory environments. We demonstrate the power of this system by collecting, segmenting and analyzing over 18 hours of courtship song from 75 males from five wild-type strains of Drosophila melanogaster. Our analysis reveals previously undetected modulation of courtship song features and extensive natural genetic variation for most components of courtship song. Despite having a large dataset with sufficient power to detect subtle modulations of song, we were unable to identify previously reported periodic rhythms in the inter-pulse interval of song. We provide detailed instructions for assembling the hardware and for using our open-source segmentation software.
Conclusions: Analysis of a large dataset of acoustic signals from Drosophila melanogaster provides novel insight into the structure and dynamics of species-specific courtship songs. Our new system for recording and analyzing fly acoustic signals should therefore greatly accelerate future studies of the genetics, neurobiology and evolution of courtship song.
Figures
Comment in
-
Can You Hear Me Now?Neuron. 2016 Feb 3;89(3):425-7. doi: 10.1016/j.neuron.2016.01.030. Neuron. 2016. PMID: 26844825
References
-
- Ewing AW, Bennet-Clark HC. The courtship songs of Drosophila. Behaviour. 1968;31:288–301. doi: 10.1163/156853968X00298. - DOI
-
- Ritchie MG, Gleason JM. Rapid evolution of courtship song pattern in Drosophila-Willistoni sibling species. J Evolution Biol. 1995;8:463–479. doi: 10.1046/j.1420-9101.1995.8040463.x. - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
