

Transcription factor Klf4, induced in the lung by oxygen at birth, regulates perinatal fibroblast and myofibroblast differentiation
- PMID: 23372771
- PMCID: PMC3553006
- DOI: 10.1371/journal.pone.0054806
Transcription factor Klf4, induced in the lung by oxygen at birth, regulates perinatal fibroblast and myofibroblast differentiation
Abstract
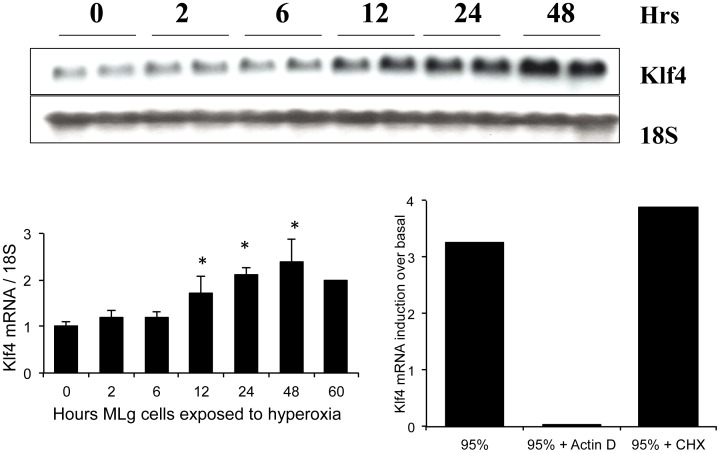
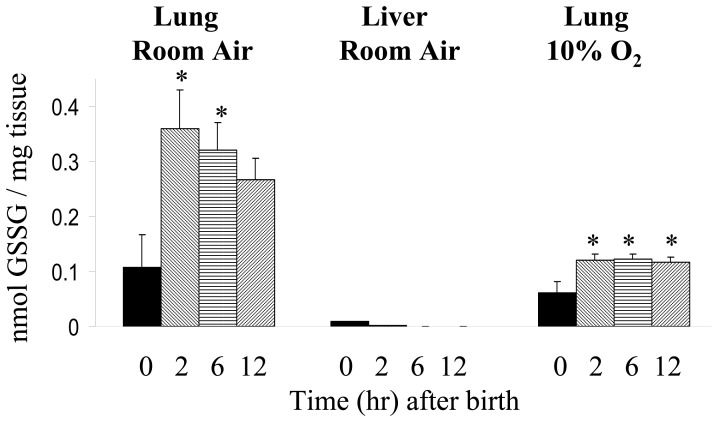
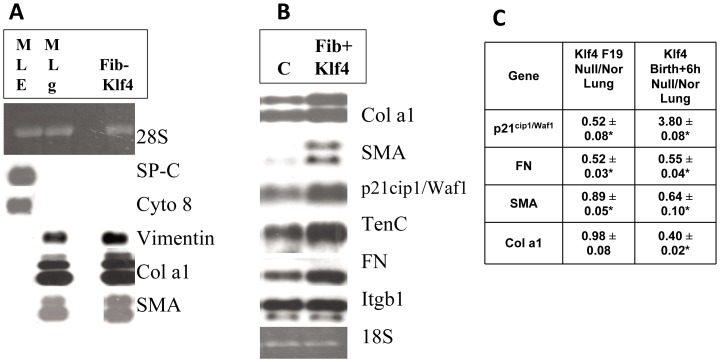
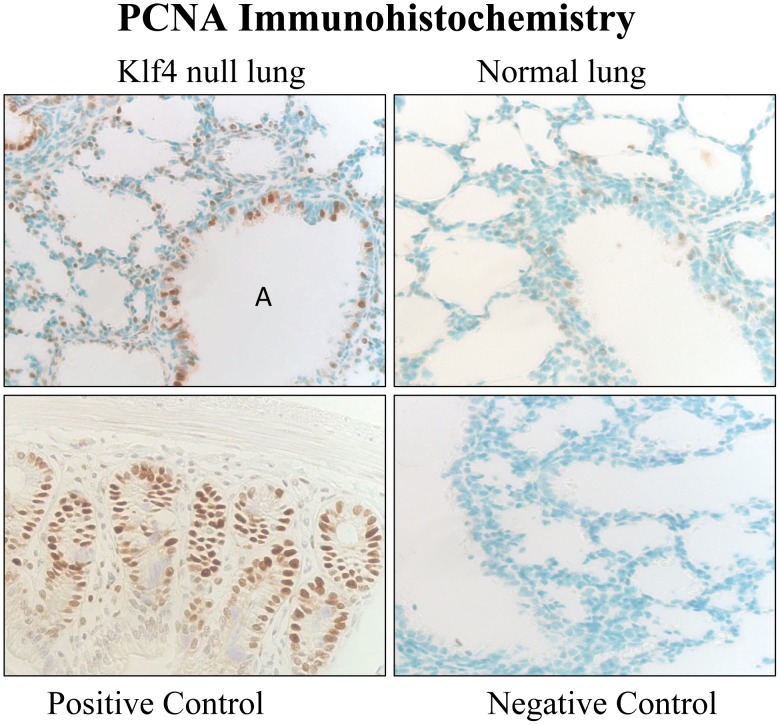
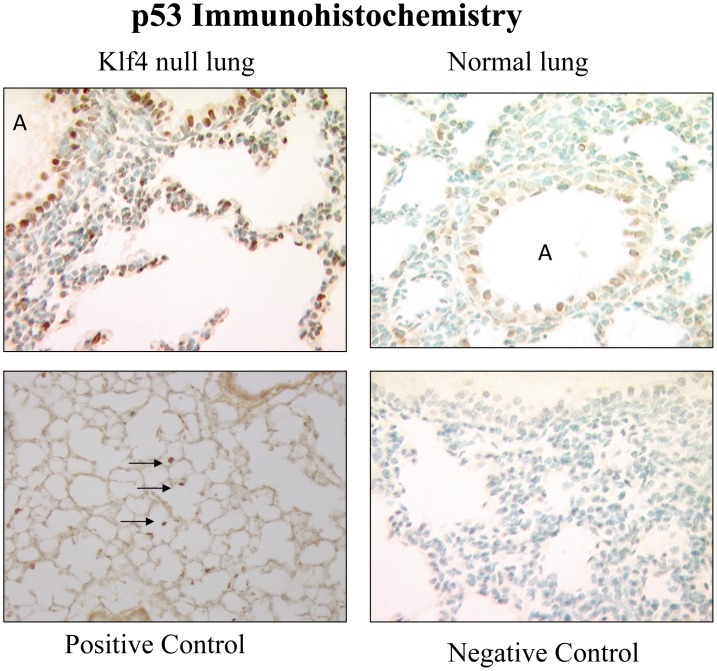
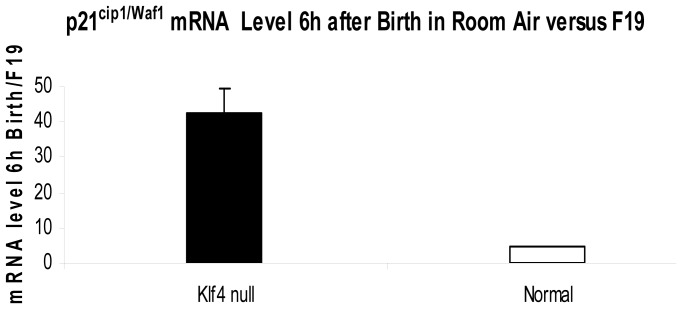
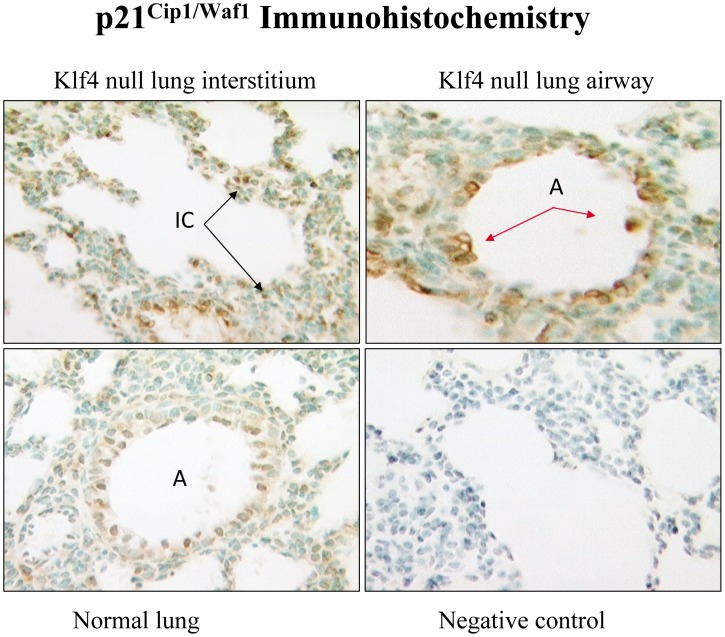
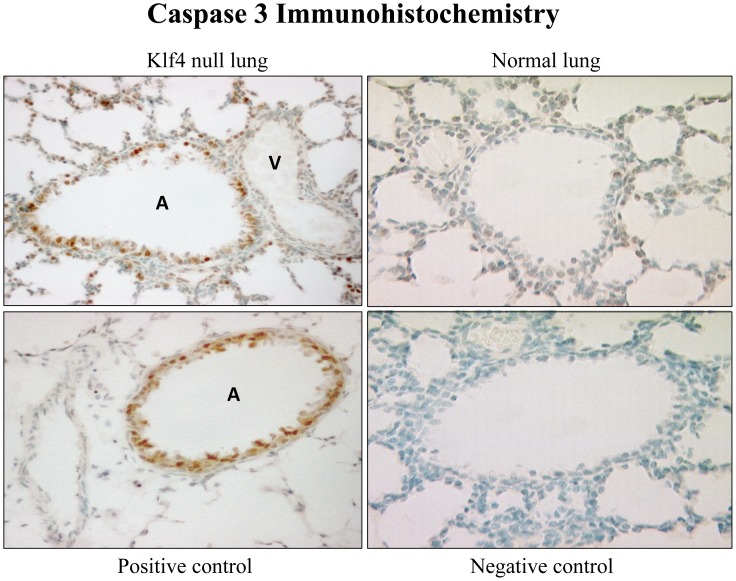
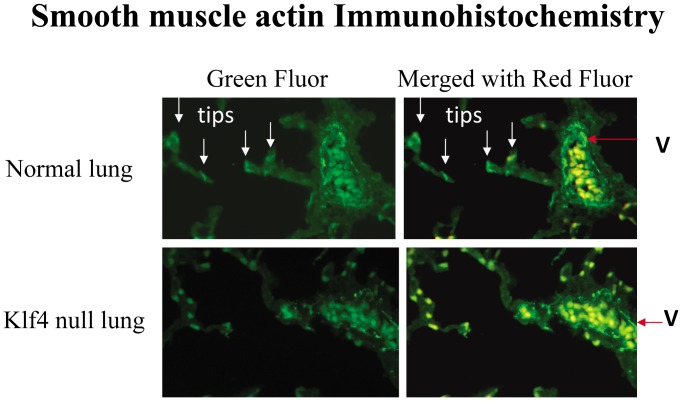
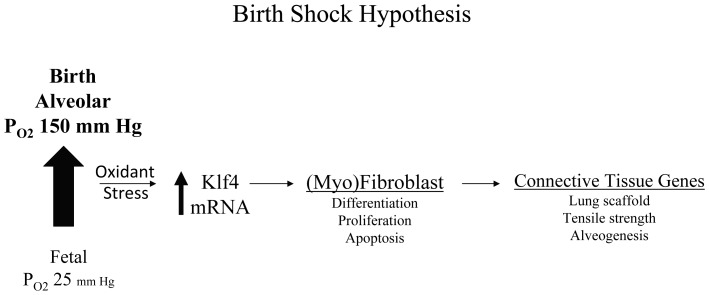
The fluid-filled lung exists in relative hypoxia in utero (∼25 mm Hg), but at birth fills with ambient air where the partial pressure of oxygen is ∼150 mm Hg. The impact of this change was studied in mouse lung with microarrays to analyze gene expression one day before, and 2, 6, 12 and 24 hours after birth into room air or 10% O(2). The expression levels of >150 genes, representing transcriptional regulation, structure, apoptosis and antioxidants were altered 2 hrs after birth in room air but blunted or absent with birth in 10% O(2). Kruppel-like factor 4 (Klf4), a regulator of cell growth arrest and differentiation, was the most significantly altered lung gene at birth. Its protein product was expressed in fibroblasts and airway epithelial cells. Klf4 mRNA was induced in lung fibroblasts exposed to hyperoxia and constitutive expression of Klf4 mRNA in Klf4-null fibroblasts induced mRNAs for p21(cip1/Waf1), smooth muscle actin, type 1 collagen, fibronectin and tenascin C. In Klf4 perinatal null lung, p21(cip1/Waf1)mRNA expression was deficient prior to birth and associated with ongoing cell proliferation after birth; connective tissue gene expression was deficient around birth and smooth muscle actin protein expression was absent from myofibroblasts at tips of developing alveoli; p53, p21(cip1/Waf1) and caspase-3 protein expression were widespread at birth suggesting excess apoptosis compared to normal lung. We propose that the changing oxygen environment at birth acts as a physiologic signal to induce lung Klf4 mRNA expression, which then regulates proliferation and apoptosis in fibroblasts and airway epithelial cells, and connective tissue gene expression and myofibroblast differentiation at the tips of developing alveoli.
Conflict of interest statement
Figures
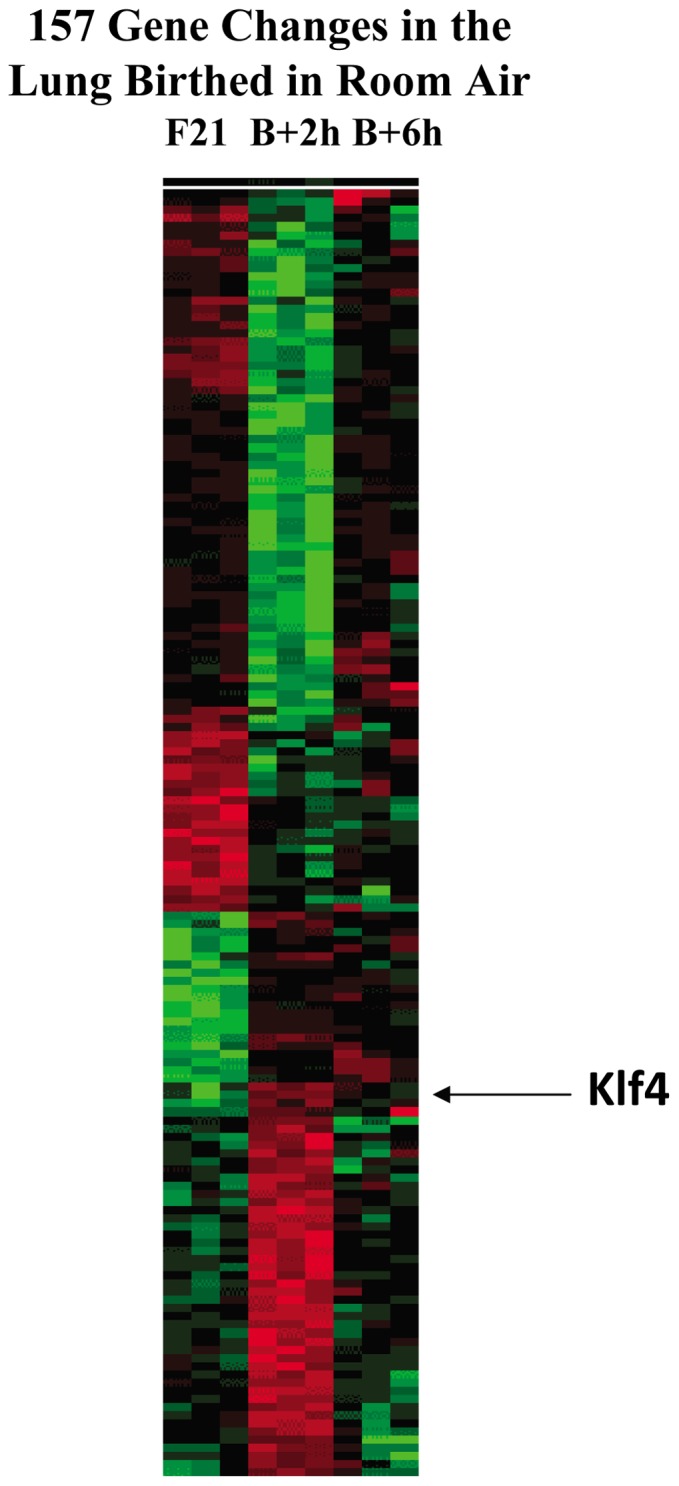
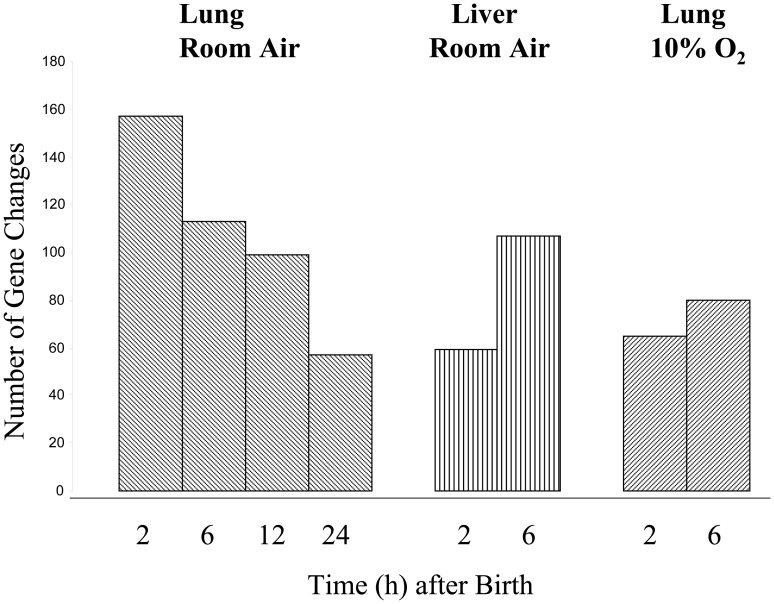
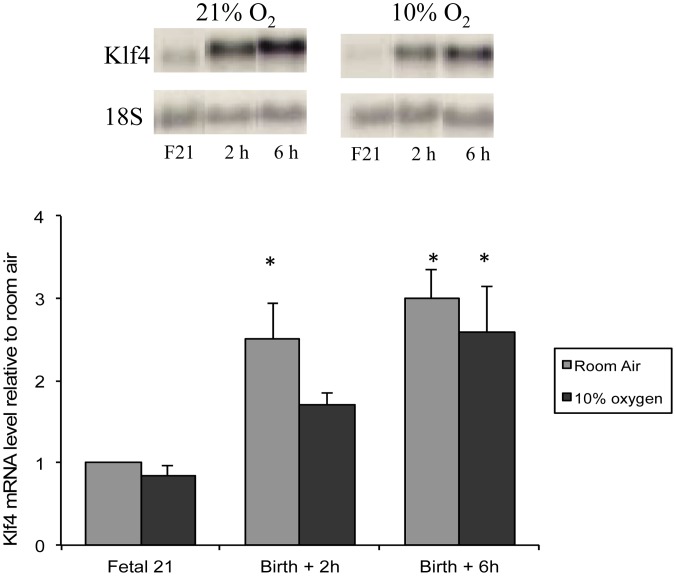
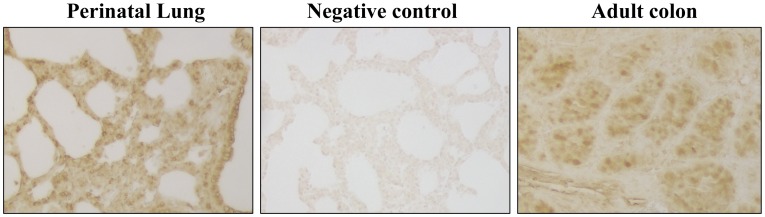
References
-
- Land SC, Wilson SM (2005) Redox regulation of lung development and perinatal lung epithelial function. Antioxid Redox Signal 7: 92–107. - PubMed
-
- Frank L, Price LT, Whitney PL (1996) Possible Mechanism for Late Gestational Development of the Antioxidant Enzymes in the Fetal Rat Lung. Biol Neonate 70: 116–27. - PubMed
-
- Chen Y, Frank L (1993) Differential gene expression of antioxidant enzymes in the perinatal rat lung. Pediatric Research 34: 27–31. - PubMed
-
- Oakes SM, Takahashi Y, Williams MC, Joyce-Brady M (1997) Ontogeny of gamma-glutamyltransferase in the rat lung. Am J Physiol 272: L739–L744. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
