

Review
doi: 10.1021/bi301504m.
Epub 2013 Feb 12.
Importance of protein dynamics during enzymatic C-H bond cleavage catalysis
Affiliations
- PMID: 23373460
- PMCID: PMC4091948
- DOI: 10.1021/bi301504m
Item in Clipboard
Review
Importance of protein dynamics during enzymatic C-H bond cleavage catalysis
Biochemistry.
.
Abstract
Quantum tunneling and protein dynamics have emerged as important components of enzyme function. This review focuses on soybean lipoxygenase-1, to illustrate how the properties of enzymatic C-H bond activation link protein motions to the fundamental bond making-breaking processes.
Conflict of interest statement
The authors declare no competing financial interest.
Figures

Illustrative free energy profiles and Arrhenius plots for the cleavage of a C–L bond, where L is H, D, or T. (A) Semiclassical KIEs predict extrapolation at high temperatures to Arrhenius prefactors that are close to being independent of isotopic labeling. (B) Tunneling correction models predict a larger value for both Ea and AL in the case of the heavier isotope(s). (C) Full tunneling models are able to reproduce data where there is no or little difference in the magnitude of Ea among the isotopes and values of AL that are greatly elevated for H in relation to D and T.

Rate constant for tunneling, as formulated in ref , comprised of three exponential terms. (a) Heavy atom motions that lead to degeneracy of reactant and product wells, a prerequisite for hydrogen tunneling between the donor and acceptor. The rate of reaching this state depends on the reaction driving force (ΔG°) and the reorganization energy (λ). (b) Resulting wave function overlap that is dependent on mass (mH), frequency (ωH), and distance (rH). The wave function overlap is shown as being greater for protium than for deuterium. (c) Energy dependence (Ex) for sampling of different donor–acceptor distances (rx). The increased energy at shorter donor–acceptor distances is offset by a more efficient wave function overlap. Adapted from ref .

Model to illustrate the impact of active site mutations in ht-ADH above and below the transition temperature at 30 °C. State 1 is the ideal state and represents the WT protein above 30 °C. The protein is optimally flexible (in the black region), and this generates active site compression that is accompanied by a close approach of the H-donor and H-acceptor. State 2 represents the situation following active site mutation above 30 °C. The white region, representing the active site, is shown in an artificially enlarged manner to allow a depiction of the resulting increased distance between the H-donor and H-acceptor that is accompanied by a decrease in the force constant for DAD sampling. State 3 is the situation that results from the combination of active site mutation and low temperature. Once again, the region representing the active site (white) is enlarged, to allow depiction of the increase in the donor–acceptor distance. Under the conditions of state 3, the increased rigidity of the protein below 30 °C prevents any “recovery” via DAD sampling and the KIE once again becomes temperature-independent.
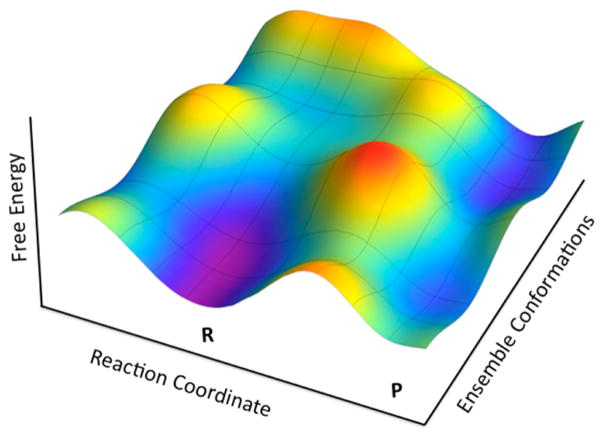
Dependence of the chemical coordinate (Reaction Coordinate) on the conformational substates (Ensemble Conformations). As illustrated, each conformational substate converts substrate to product with a distinctive rate constant (provided by R. Larsen and A. Kohen). A similar picture has been introduced by Benkovic et al. to illustrate the role of a conformational landscape in enzyme catalysis.

X-ray structure of SLO-1, with LA modeled into the active site.
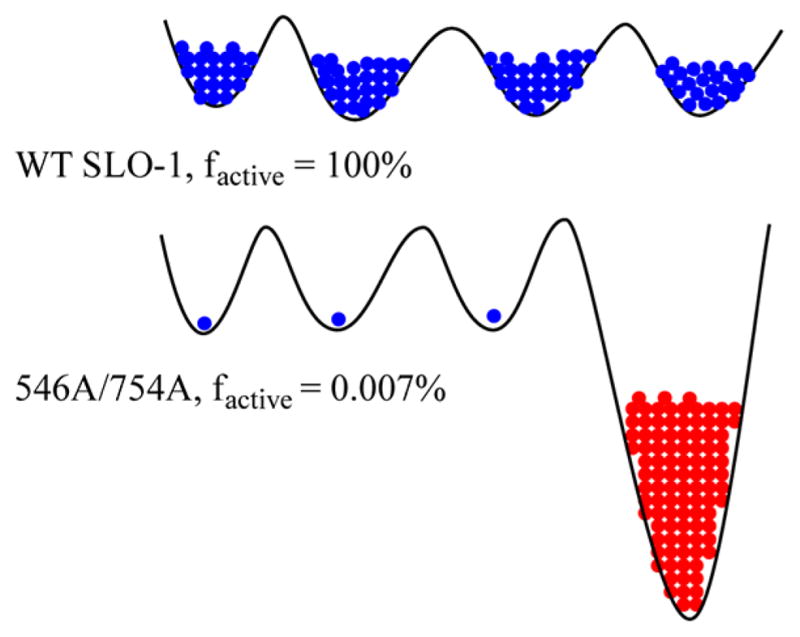
Schematic impact of deleterious mutation on the conformational landscape of SLO-1. The conformational landscape of the enzyme is shown by solid black lines with energy increasing along the vertical axis. Blue indicates active conformers and red inactive conformer(s). In the case of WT SLO-1, the enzyme molecules are fully populated in the catalytically active regions of the conformational spaces. In the double mutant, Leu546Ala/Leu754Ala, only a small proportion of the enzyme is populated in the catalytically active region of the conformational space and a significant proportion of enzyme occupies deep local minima of catalytically very slow and/or inactive conformations. For the sake of simplicity, only one deep local minimum is shown. Different conformational landscapes are expected for the free enzyme (controlling kcat/Km(LA)) and the E·S′ complex (controlling kcat). In the example shown, the fraction of active enzyme is obtained from the relative kcat numbers.


aThe standard state for the binding of substrate has been arbitrarily set at the Kd for substrate, resulting in equal energies for E+S and E·S.

a(A) Reaction of [11-H2]LA, corrected for kinetic complexity. (B) Reaction of [11-2H2]LA. (C) Computed KIEs using an optimized structure for LA bound to SLO-1 and the assumption of a freely equilibrated pentadienyl radical intermediate (cf. Scheme 1).
Similar articles
-
Active Site Dynamical Effects in the Hydrogen Transfer Rate-limiting Step in the Catalysis of Linoleic Acid by Soybean Lipoxygenase-1 (SLO-1): Primary and Secondary Isotope Contributions.J Phys Chem B. 2015 Jul 30;119(30):9532-46. doi: 10.1021/acs.jpcb.5b02385. Epub 2015 Jul 14. J Phys Chem B. 2015. PMID: 26079999
-
Catalytic efficiency of enzymes: a theoretical analysis.Biochemistry. 2013 Mar 26;52(12):2012-20. doi: 10.1021/bi301515j. Epub 2012 Dec 20. Biochemistry. 2013. PMID: 23240765 Free PMC article. Review.
-
Environmentally coupled hydrogen tunneling. Linking catalysis to dynamics.Eur J Biochem. 2002 Jul;269(13):3113-21. doi: 10.1046/j.1432-1033.2002.03022.x. Eur J Biochem. 2002. PMID: 12084051 Review.
-
Nature of hydrogen transfer in soybean lipoxygenase 1: separation of primary and secondary isotope effects.Biochemistry. 1999 Sep 21;38(38):12218-28. doi: 10.1021/bi990834y. Biochemistry. 1999. PMID: 10493789
-
Temperature-dependent isotope effects in soybean lipoxygenase-1: correlating hydrogen tunneling with protein dynamics.J Am Chem Soc. 2002 Apr 17;124(15):3865-74. doi: 10.1021/ja012205t. J Am Chem Soc. 2002. PMID: 11942823
Cited by
-
A Reactive Manganese(IV)-Hydroxide Complex: A Missing Intermediate in Hydrogen Atom Transfer by High-Valent Metal-Oxo Porphyrinoid Compounds.J Am Chem Soc. 2018 Mar 28;140(12):4380-4390. doi: 10.1021/jacs.8b00350. Epub 2018 Mar 15. J Am Chem Soc. 2018. PMID: 29542921 Free PMC article.
-
The Use of Multiscale Molecular Simulations in Understanding a Relationship between the Structure and Function of Biological Systems of the Brain: The Application to Monoamine Oxidase Enzymes.Front Neurosci. 2016 Jul 15;10:327. doi: 10.3389/fnins.2016.00327. eCollection 2016. Front Neurosci. 2016. PMID: 27471444 Free PMC article. Review.
-
Mimicking Elementary Reactions of Manganese Lipoxygenase Using Mn-hydroxo and Mn-alkylperoxo Complexes.Molecules. 2021 Nov 25;26(23):7151. doi: 10.3390/molecules26237151. Molecules. 2021. PMID: 34885729 Free PMC article.
-
Reflections on the catalytic power of a TIM-barrel.Bioorg Chem. 2014 Dec;57:206-212. doi: 10.1016/j.bioorg.2014.07.001. Epub 2014 Jul 11. Bioorg Chem. 2014. PMID: 25092608 Free PMC article.
-
Dynamical network of residue-residue contacts reveals coupled allosteric effects in recognition, catalysis, and mutation.Proc Natl Acad Sci U S A. 2016 Apr 26;113(17):4735-40. doi: 10.1073/pnas.1523573113. Epub 2016 Apr 11. Proc Natl Acad Sci U S A. 2016. PMID: 27071107 Free PMC article.
References
-
- Pauling L. Chemical achievements and hope for the future. Am Sci. 1948;36:51–58. - PubMed
-
- Hilvert D. Critical analysis of antibody catalysis. Annu Rev Biochem. 2000;69:751–793. - PubMed
-
- Rothlisberger D, Khersonsky O, Wollacott AM, Jiang L, DeChancie J, Betker J, Gallaher JL, Althoff EA, Zanghellini A, Dym O, Albeck S, Houk KN, Tawfik DS, Baker D. Kemp elimination catalysts by computational enzyme design. Nature. 2008;453:190–195. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
