

The PB2, PA, HA, NP, and NS genes of a highly pathogenic avian influenza virus A/whooper swan/Mongolia/3/2005 (H5N1) are responsible for pathogenicity in ducks
- PMID: 23374292
- PMCID: PMC3570374
- DOI: 10.1186/1743-422X-10-45
The PB2, PA, HA, NP, and NS genes of a highly pathogenic avian influenza virus A/whooper swan/Mongolia/3/2005 (H5N1) are responsible for pathogenicity in ducks
Abstract
Background: Wild ducks are the natural hosts of influenza A viruses. Duck influenza, therefore, has been believed inapparent infection with influenza A viruses, including highly pathogenic avian influenza viruses (HPAIVs) in chickens. In fact, ducks experimentally infected with an HPAIV strain, A/Hong Kong/483/1997 (H5N1) (HK483), did not show any clinical signs. Another HPAIV strain, A/whooper swan/Mongolia/3/2005 (H5N1) (MON3) isolated from a dead swan, however, caused neurological dysfunction and death in ducks.
Method: To understand the mechanism whereby MON3 shows high pathogenicity in ducks, HK483, MON3, and twenty-four reassortants generated between these two H5N1 viruses were compared for their pathogenicity in domestic ducks.
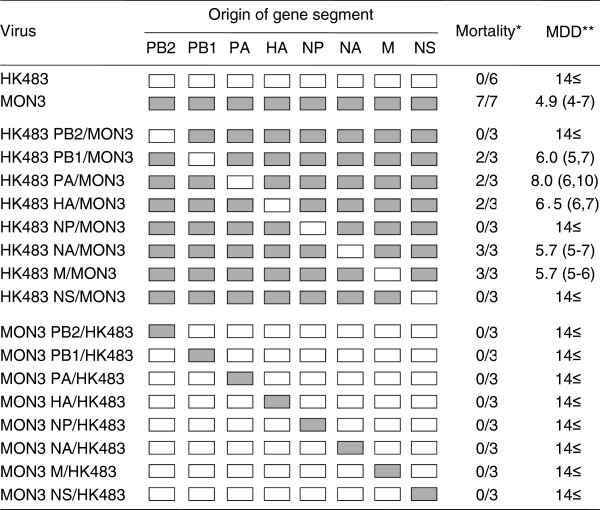
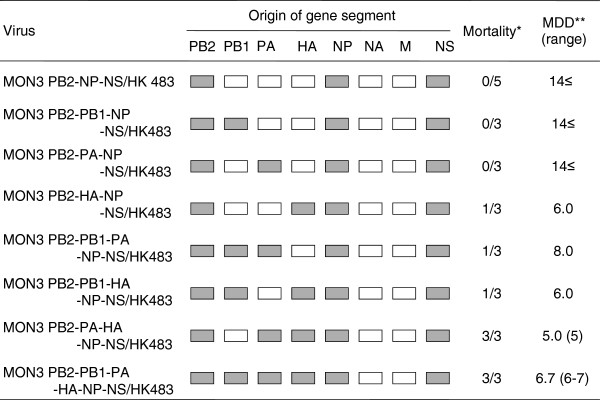
Results: None of the ducks infected with MON3-based single-gene reassortants bearing the PB2, NP, or NS gene segment of HK483 died, and HK483-based single-gene reassortants bearing PB2, NP, or NS genes of MON3 were not pathogenic in ducks, suggesting that multiple gene segments contribute to the pathogenicity of MON3 in ducks. All the ducks infected with the reassortant bearing PB2, PA, HA, NP, and NS gene segments of MON3 died within five days post-inoculation, as did those infected with MON3. Each of the viruses was assessed for replication in ducks three days post-inoculation. MON3 and multi-gene reassortants pathogenic in ducks were recovered from all of the tissues examined and replicated with high titers in the brains and lungs.
Conclusion: The present results indicate that multigenic factors are responsible for efficient replication of MON3 in ducks. In particular, virus growth in the brain might correlate with neurological dysfunction and the disease severity.
Figures
References
-
- Kawaoka Y, Cox NJ, Haller O, Hongo S, Kaverin N, Klenk HD, Lamb RA, McCauley J, Palese P, Rimstad E, Webster RG. In: Virus Taxonomy. 8. Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA, editor. California: Elsevier Academic Press; 2005. Family Orthomyxoviridae; pp. 681–693.
-
- Fouchier RA, Munster V, Wallensten A, Bestebroer TM, Herfst S, Smith D, Rimmelzwaan GF, Olsen B, Osterhaus AD. Characterization of a novel influenza A virus hemagglutinin subtype (H16) obtained from black-headed gulls. J Virol. 2005;79:2814–2822. doi: 10.1128/JVI.79.5.2814-2822.2005. - DOI - PMC - PubMed
-
- Okazaki K, Takada A, Ito T, Imai M, Takakuwa H, Hatta M, Ozaki H, Tanizaki T, Nagano T, Ninomiya A, Demenov VA, Tyaptirganov MM, Karatayeva TD, Yamnikova SS, Lvov DK, Kida H. Precursor genes of future pandemic influenza viruses are perpetuated in ducks nesting in Siberia. Arch Virol. 2000;145:885–893. doi: 10.1007/s007050050681. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
