

Influence of GluN2 subunit identity on NMDA receptor function
- PMID: 23376022
- PMCID: PMC3778433
- DOI: 10.1016/j.neuropharm.2013.01.016
Influence of GluN2 subunit identity on NMDA receptor function
Abstract
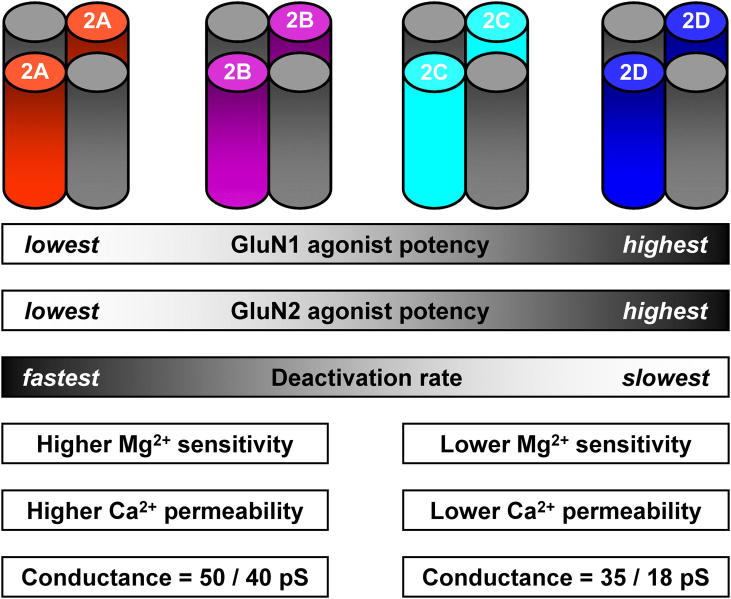
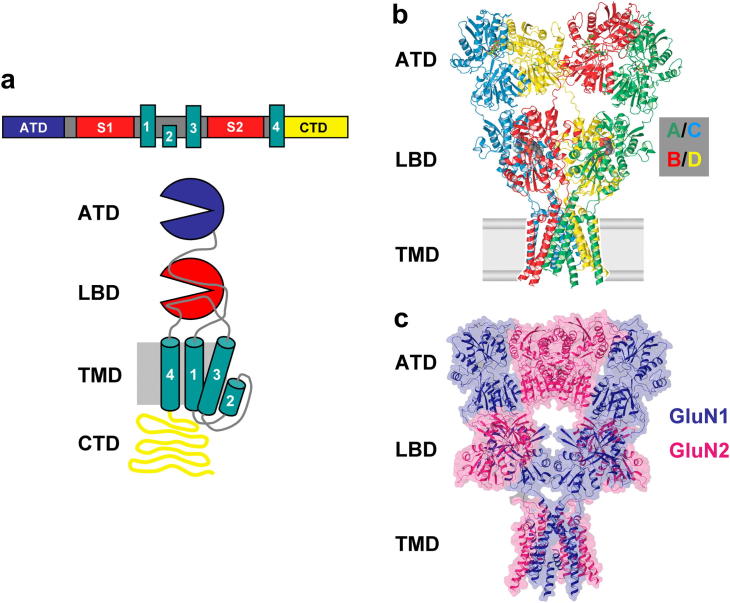
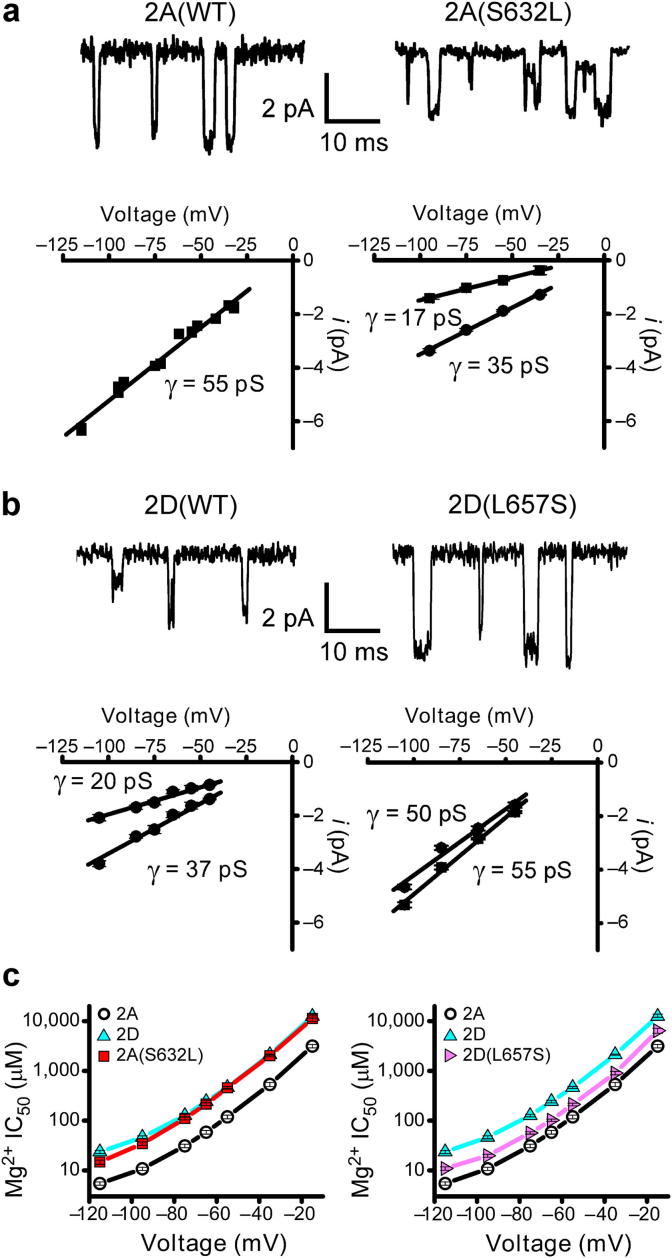
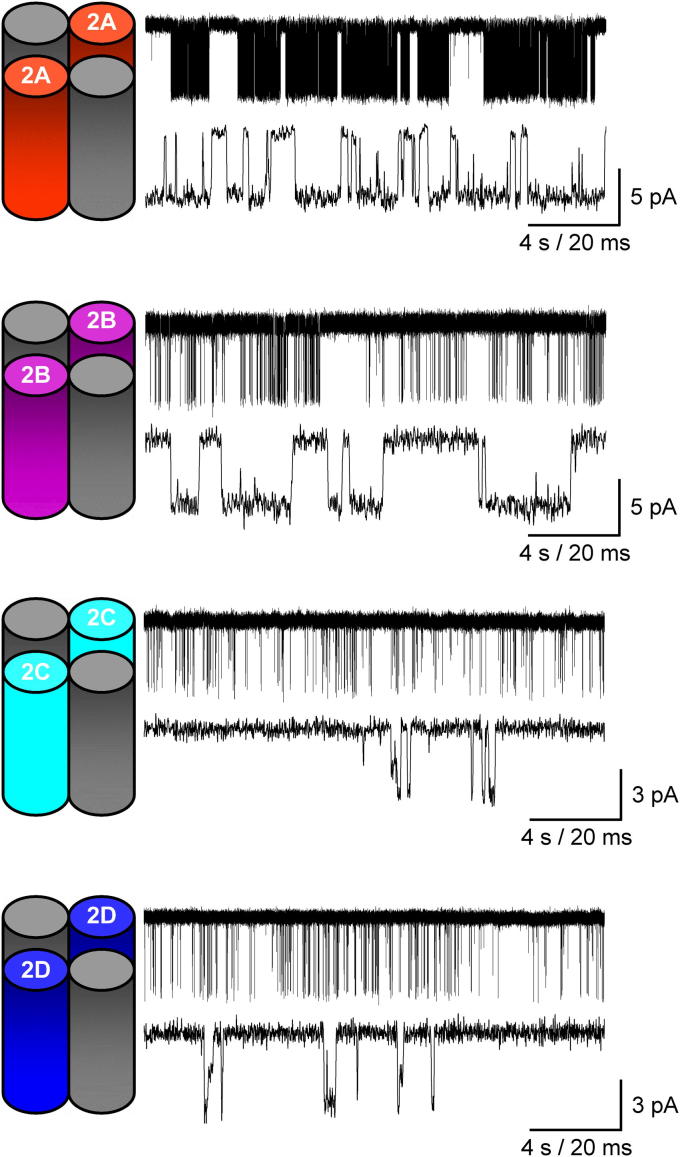
N-methyl-d-aspartate receptors (NMDARs) are ligand-gated ion channels ('ionotropic' receptors) activated by the major excitatory neurotransmitter, l-glutamate. While the term 'the NMDAR' is often used it obscures the fact that this class of receptor contains within it members whose properties are as different as they are similar. This heterogeneity was evident from early electrophysiological, pharmacological and biochemical assessments of the functional properties of NMDARs and while the molecular basis of this heterogeneity has taken many years to elucidate, it indicated from the outset that the diversity of NMDAR phenotypes could allow this receptor family to subserve a variety of functions in the mammalian central nervous system. In this review we highlight some recent studies that have identified structural elements within GluN2 subunits that contribute to the heterogeneous biophysical properties of NMDARs, consider why some recently described novel pharmacological tools may permit better identification of native NMDAR subtypes, examine the evidence that NMDAR subtypes differentially contribute to the induction of long-term potentiation and long-term depression and discuss how through the use of chimeric proteins additional insights have been obtained that account for NMDAR subtype-dependency of physiological and pathophysiological signalling. This article is part of the Special Issue entitled 'Glutamate Receptor-Dependent Synaptic Plasticity'.
Keywords: Biophysics; Excitoxicity; Glutamate receptor; NMDA receptor; Pharmacology; Plasticity; Structure.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
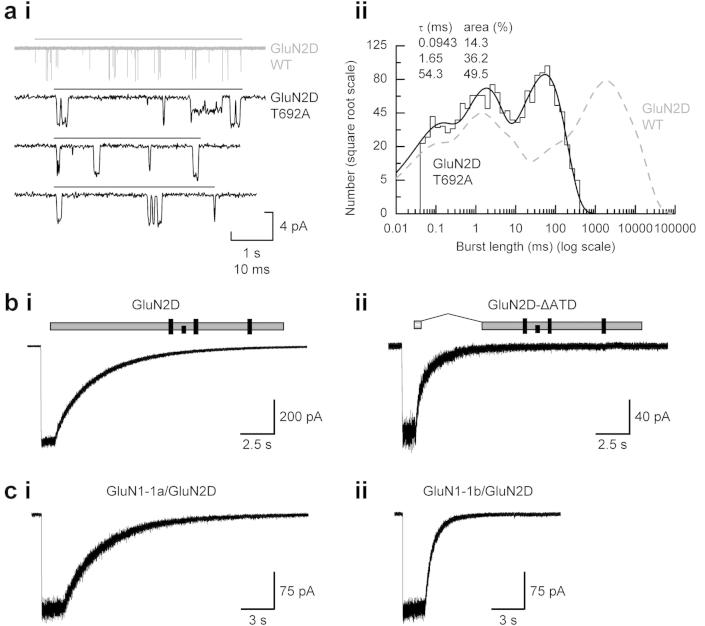
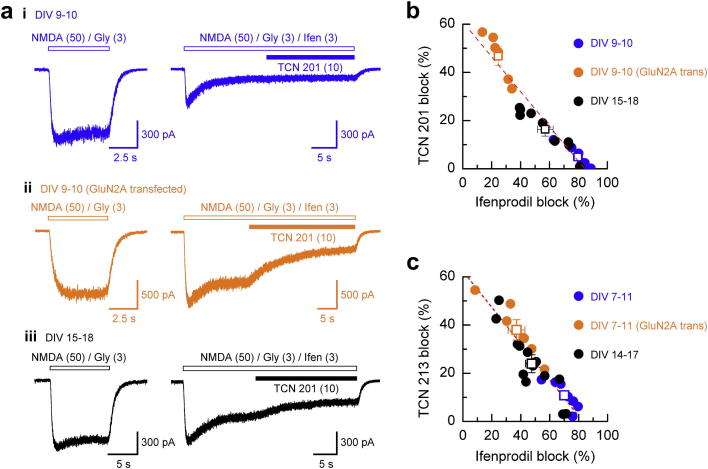
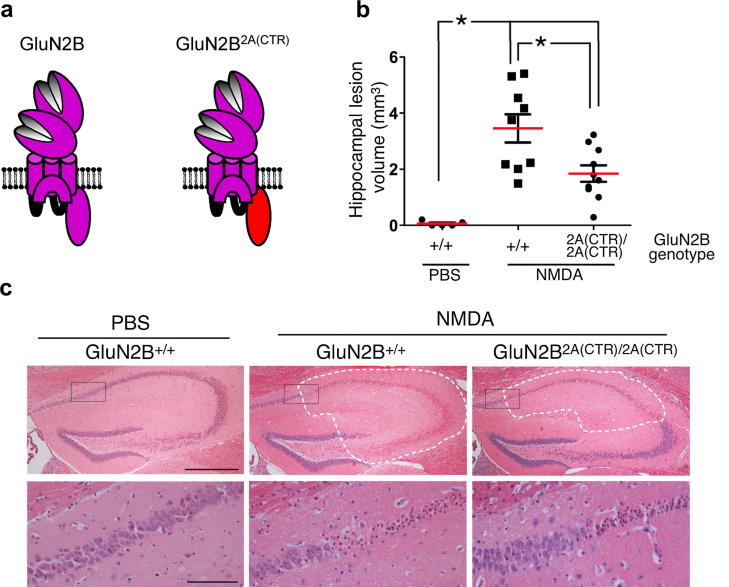
References
-
- Aarts M., Liu Y., Liu L., Besshoh S., Arundine M., Gurd J.W., Wang Y.T., Salter M.W., Tymianski M. Treatment of ischemic brain damage by perturbing NMDA receptor- PSD-95 protein interactions. Science. 2002;298:846–850. - PubMed
-
- Acker T.M., Yuan H., Hansen K.B., Vance K.M., Ogden K.K., Jensen H.S., Burger P.B., Mullasseril P., Snyder J.P., Liotta D.C., Traynelis S.F. Mechanism for noncompetitive inhibition by novel GluN2C/D N-methyl-d-aspartate receptor subunit-selective modulators. Mol. Pharmacol. 2011;80:782–795. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
