

Oncogenic Ras suppresses Cdk1 in a complex manner during the incubation of activated Xenopus egg extracts
- PMID: 23376039
- PMCID: PMC3809123
- DOI: 10.1016/j.abb.2013.01.006
Oncogenic Ras suppresses Cdk1 in a complex manner during the incubation of activated Xenopus egg extracts
Abstract
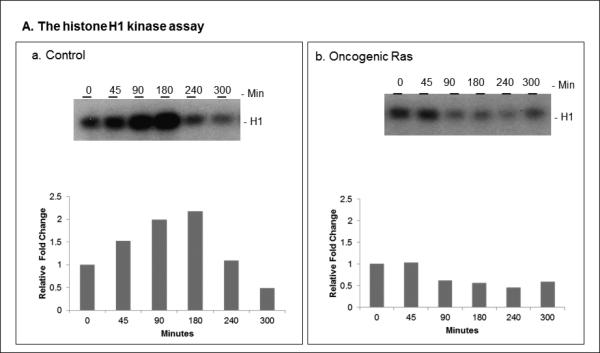
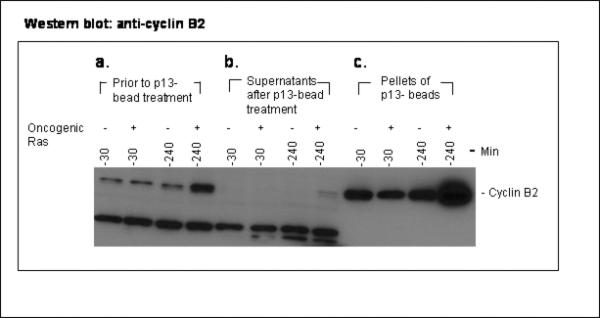
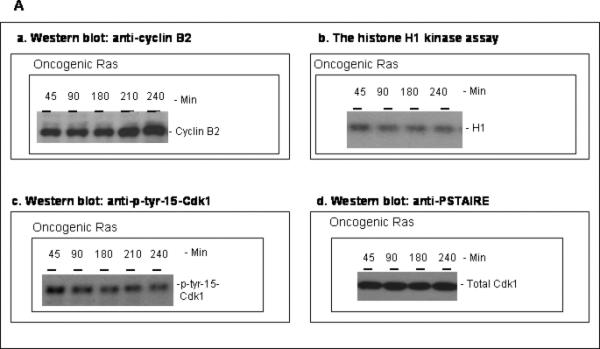
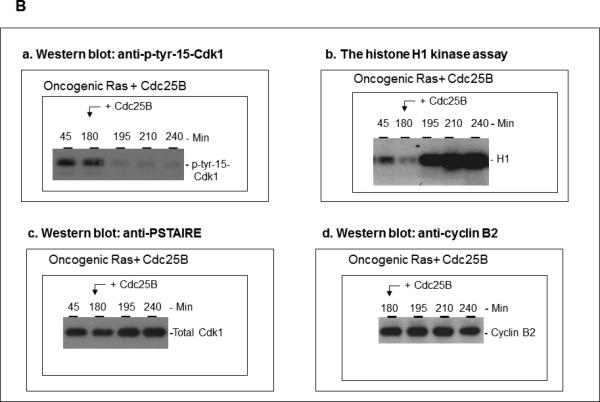
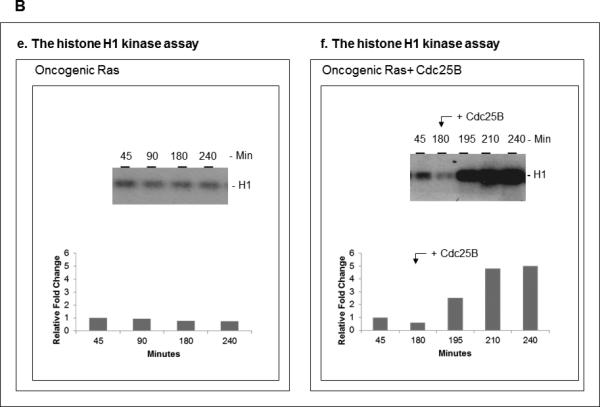
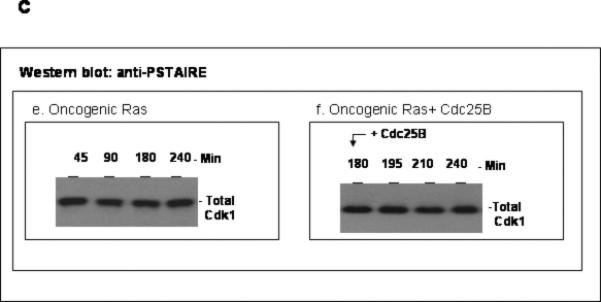
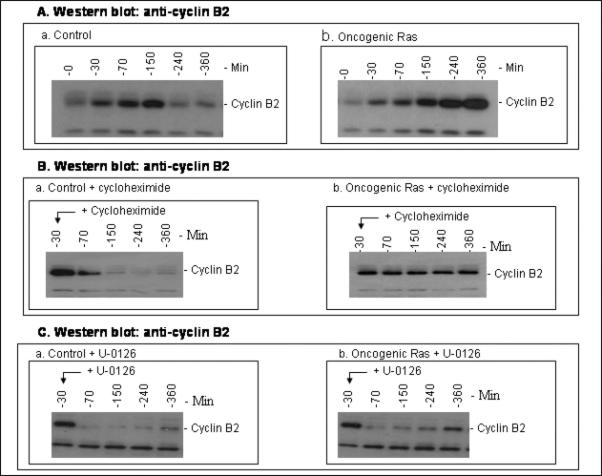
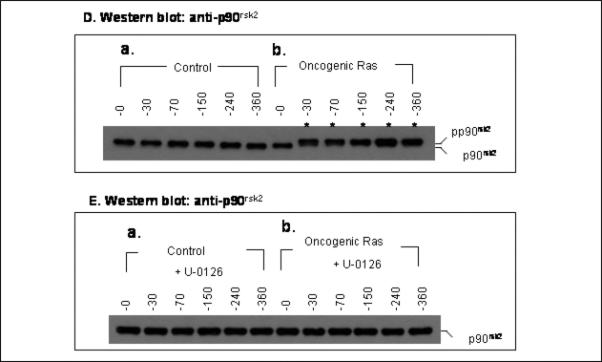
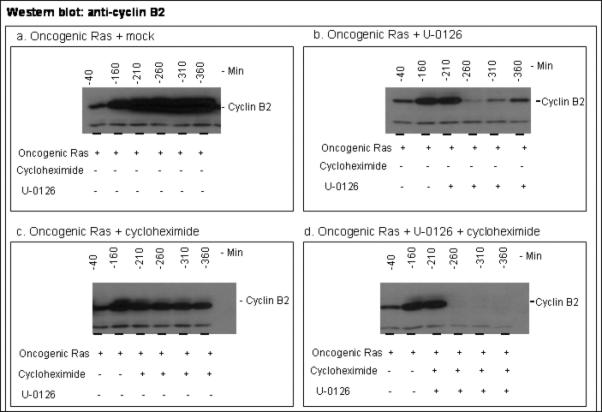
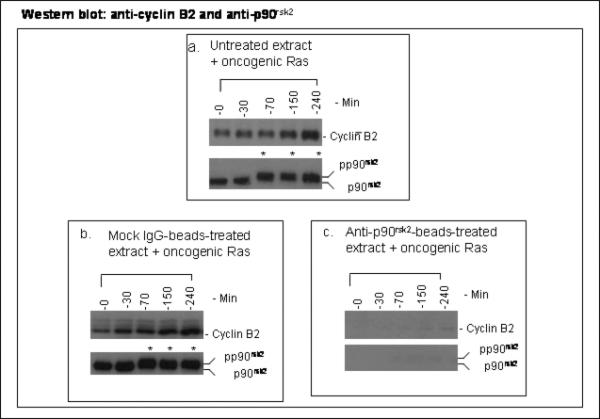
The activity of Cdk1 is the driving force for entry into M-phase during the cell cycle. Activation of Cdk1 requires synthesis and accumulation of cyclin B, binding of cyclin B to Cdk1, and removal of the inhibitory tyr-15-Cdk1 phosphorylation. It was previously shown that oncogenic Ras suppresses Cdk1 activation during the incubation of activated Xenopus egg extracts. However, how oncogenic Ras suppresses Cdk1 remained unclear. Using the histone H1 kinase assay to follow Cdk1 activity and Western blot analysis to assess levels of both cyclin B2 and phosphorylated-tyr-15-Cdk1, how oncogenic Ras suppresses Cdk1 is studied. The results indicate that oncogenic Ras suppresses Cdk1 via induction of persistent phosphorylation of tyr-15-Cdk1. Interestingly, the results reveal that, compared with cyclin B2 in control activated egg extracts, which increased, peaked and then declined during the incubation, oncogenic Ras induced continuous accumulation of cyclin B2. The results also indicate that oncogenic Ras induces continuous accumulation of cyclin B2 primarily through stabilization of cyclin B2, which is mediated by constitutive activation of the Raf-Mek-Erk-p90(rsk) pathway. Taken together, these results indicate that oncogenic Ras suppresses Cdk1 in a complex manner: It induces continuous accumulation of cyclin B2, but also causes persistent inhibitory phosphorylation of tyr-15-Cdk1.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures


Similar articles
-
A novel role for Cdk1/cyclin B in regulating B-raf activation at mitosis.Mol Biol Cell. 2008 Jul;19(7):2907-15. doi: 10.1091/mbc.e07-07-0679. Epub 2008 Apr 23. Mol Biol Cell. 2008. PMID: 18434602 Free PMC article.
-
Cyclin B2/cyclin-dependent kinase1 dissociation precedes CDK1 Thr-161 dephosphorylation upon M-phase promoting factor inactivation in Xenopus laevis cell-free extract.Int J Dev Biol. 2007;51(4):297-305. doi: 10.1387/ijdb.072292fc. Int J Dev Biol. 2007. PMID: 17554681
-
Cyclin B dissociation from CDK1 precedes its degradation upon MPF inactivation in mitotic extracts of Xenopus laevis embryos.Cell Cycle. 2006 Aug;5(15):1687-98. doi: 10.4161/cc.5.15.3123. Epub 2006 Aug 1. Cell Cycle. 2006. PMID: 16921258
-
A link between MAP kinase and p34(cdc2)/cyclin B during oocyte maturation: p90(rsk) phosphorylates and inactivates the p34(cdc2) inhibitory kinase Myt1.EMBO J. 1998 Sep 1;17(17):5037-47. doi: 10.1093/emboj/17.17.5037. EMBO J. 1998. PMID: 9724639 Free PMC article.
-
The 96 kDa protein kinase activated by oncogenic Ras in Xenopus egg extracts is also activated by constitutively active Mek: activation requires serine/threonine phosphorylation.Oncogene. 1997 Apr 10;14(14):1653-60. doi: 10.1038/sj.onc.1201009. Oncogene. 1997. PMID: 9135066
Cited by
-
NCAPG-mediated CDK1 promotes malignant progression of non-small cell lung cancer via ERK signaling activation.Am J Cancer Res. 2024 Nov 15;14(11):5338-5350. doi: 10.62347/DNAQ7105. eCollection 2024. Am J Cancer Res. 2024. PMID: 39659935 Free PMC article.
-
CDK1 promotes the proliferation of melanocytes in Rex rabbits.Genes Genomics. 2022 Oct;44(10):1191-1199. doi: 10.1007/s13258-022-01283-4. Epub 2022 Aug 11. Genes Genomics. 2022. PMID: 35951158
References
-
- Malumbres M, Barbacid M. RAS oncogenes: the first 30 years. Nat Rev Cancer. 2003;3:459–465. - PubMed
-
- Land H, Parada LF, Weinberg RA. Tumorigenic conversion of primary embryo fibroblasts requires at least two cooperating oncogenes. Nature. 1983;304:596–602. - PubMed
-
- Lloyd AC. Ras versus cyclin-dependent kinase inhibitors. Curr Opin Genet Dev. 1988;8:43–48. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
