

Functional dissection of Toxoplasma gondii perforin-like protein 1 reveals a dual domain mode of membrane binding for cytolysis and parasite egress
- PMID: 23376275
- PMCID: PMC3605689
- DOI: 10.1074/jbc.M113.450932
Functional dissection of Toxoplasma gondii perforin-like protein 1 reveals a dual domain mode of membrane binding for cytolysis and parasite egress
Abstract
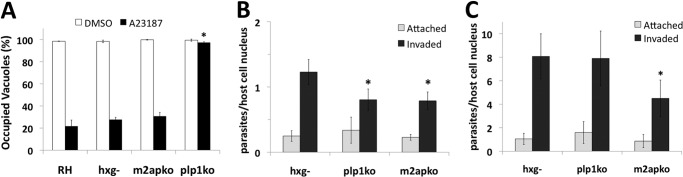
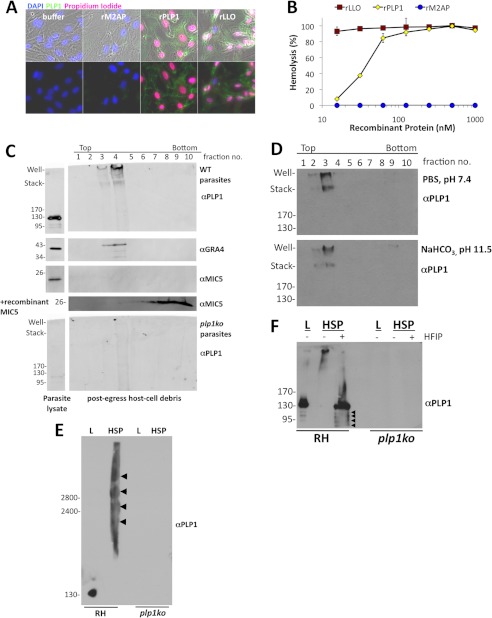
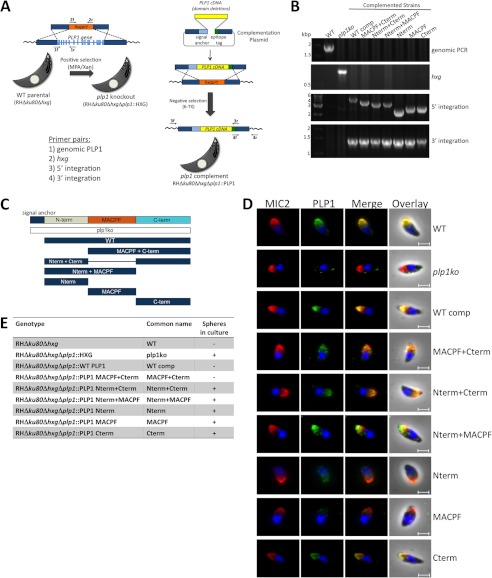
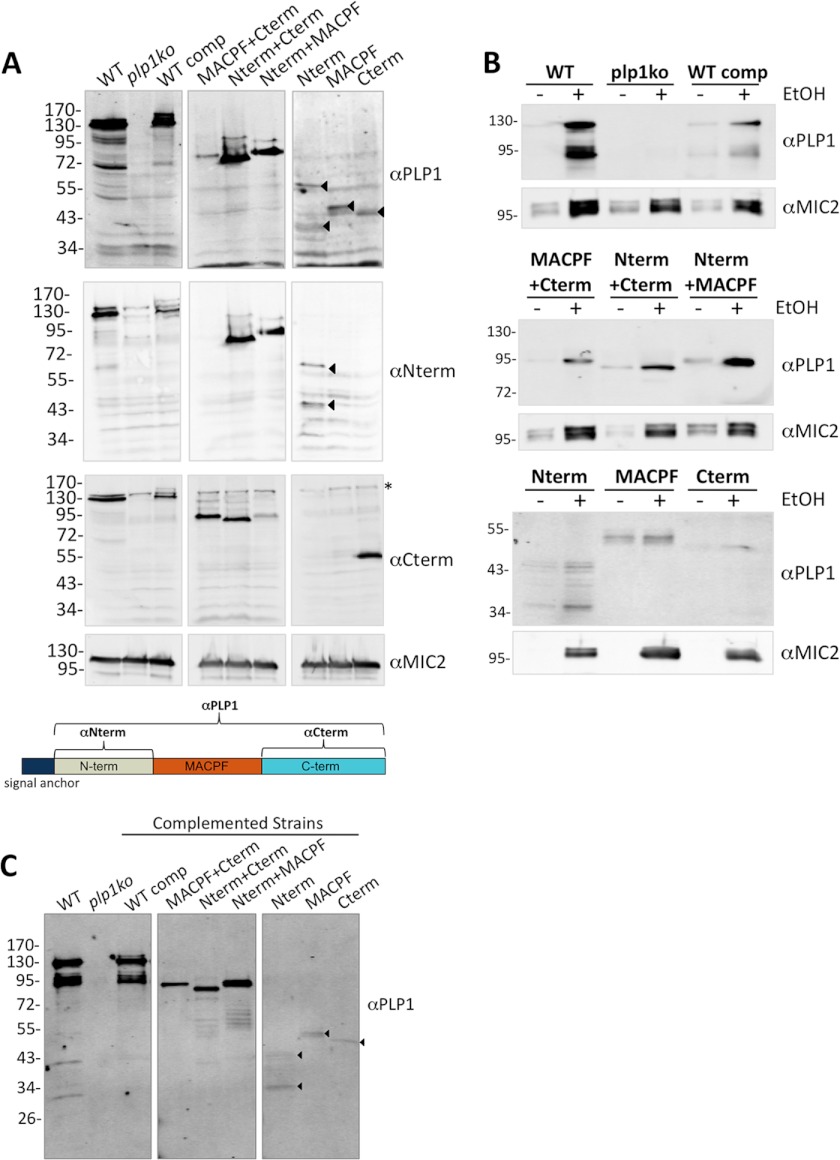
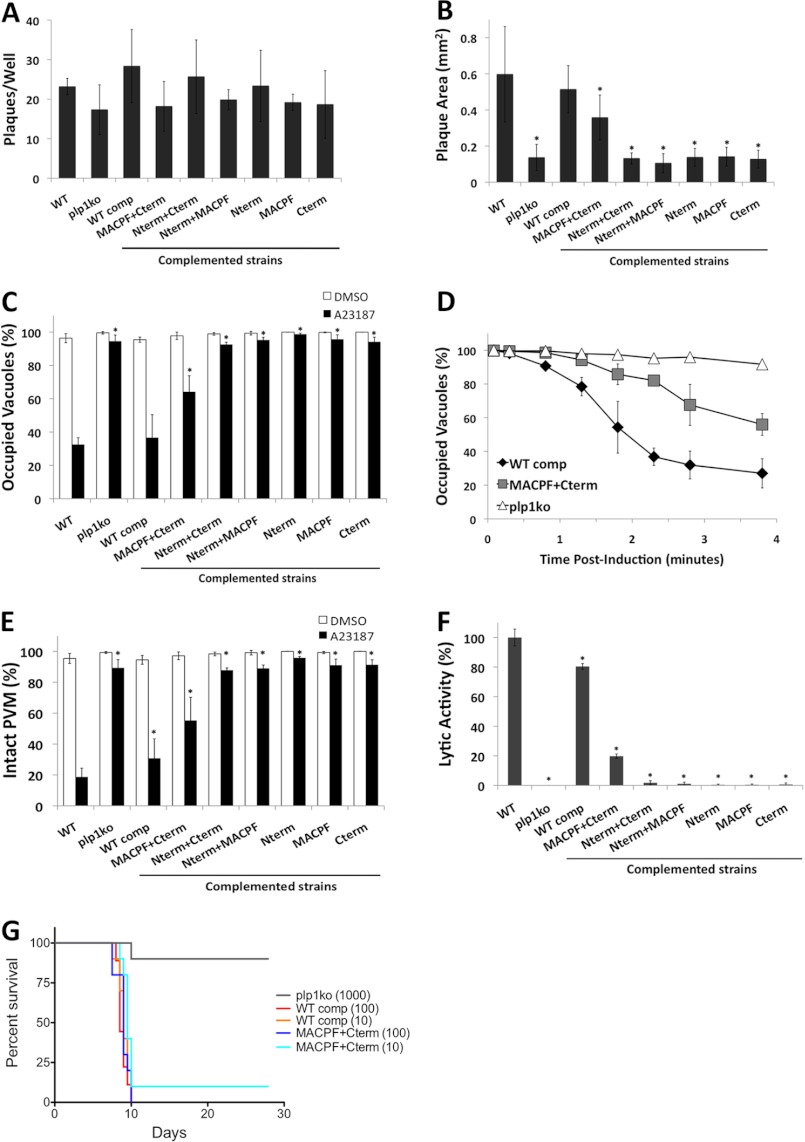
The recently discovered role of a perforin-like protein (PLP1) for rapid host cell egress by the protozoan parasite Toxoplasma gondii expanded the functional diversity of pore-forming proteins. Whereas PLP1 was found to be necessary for rapid egress and pathogenesis, the sufficiency for and mechanism of membrane attack were yet unknown. Here we further dissected the PLP1 knock-out phenotype, the mechanism of PLP1 pore formation, and the role of each domain by genetic complementation. We found that PLP1 is sufficient for membrane disruption and has a conserved mechanism of pore formation through target membrane binding and oligomerization to form large, multimeric membrane-embedded complexes. The highly conserved, central MACPF domain and the β-sheet-rich C-terminal domain were required for activity. Loss of the unique N-terminal extension reduced lytic activity and led to a delay in rapid egress, but did not significantly decrease virulence, suggesting that small amounts of lytic activity are sufficient for pathogenesis. We found that both N- and C-terminal domains have membrane binding activity, with the C-terminal domain being critical for function. This dual mode of membrane association may promote PLP1 activity and parasite egress in the diverse cell types in which this parasite replicates.
Figures

Similar articles
-
Perforin-Like Proteins of Apicomplexan Parasites.Front Cell Infect Microbiol. 2020 Sep 15;10:578883. doi: 10.3389/fcimb.2020.578883. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 33042876 Free PMC article. Review.
-
Rapid membrane disruption by a perforin-like protein facilitates parasite exit from host cells.Science. 2009 Jan 23;323(5913):530-3. doi: 10.1126/science.1165740. Epub 2008 Dec 18. Science. 2009. PMID: 19095897 Free PMC article.
-
Structural basis of Toxoplasma gondii perforin-like protein 1 membrane interaction and activity during egress.PLoS Pathog. 2018 Dec 4;14(12):e1007476. doi: 10.1371/journal.ppat.1007476. eCollection 2018 Dec. PLoS Pathog. 2018. PMID: 30513119 Free PMC article.
-
Structures of monomeric and oligomeric forms of the Toxoplasma gondii perforin-like protein 1.Sci Adv. 2018 Mar 21;4(3):eaaq0762. doi: 10.1126/sciadv.aaq0762. eCollection 2018 Mar. Sci Adv. 2018. PMID: 29750191 Free PMC article.
-
Cholesterol-dependent cytolysins.Adv Exp Med Biol. 2010;677:56-66. doi: 10.1007/978-1-4419-6327-7_5. Adv Exp Med Biol. 2010. PMID: 20687480 Review.
Cited by
-
The Apicomplexan CDC/MACPF-like pore-forming proteins.Curr Opin Microbiol. 2015 Aug;26:48-52. doi: 10.1016/j.mib.2015.05.001. Epub 2015 May 27. Curr Opin Microbiol. 2015. PMID: 26025132 Free PMC article. Review.
-
Research advances in microneme protein 3 of Toxoplasma gondii.Parasit Vectors. 2015 Jul 22;8:384. doi: 10.1186/s13071-015-1001-4. Parasit Vectors. 2015. PMID: 26194005 Free PMC article. Review.
-
Regulation of phosphoinositide metabolism in Apicomplexan parasites.Front Cell Dev Biol. 2023 Sep 15;11:1163574. doi: 10.3389/fcell.2023.1163574. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37791074 Free PMC article. Review.
-
Perforin-Like Proteins of Apicomplexan Parasites.Front Cell Infect Microbiol. 2020 Sep 15;10:578883. doi: 10.3389/fcimb.2020.578883. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 33042876 Free PMC article. Review.
-
Essential role of Plasmodium perforin-like protein 4 in ookinete midgut passage.PLoS One. 2018 Aug 13;13(8):e0201651. doi: 10.1371/journal.pone.0201651. eCollection 2018. PLoS One. 2018. PMID: 30102727 Free PMC article.
References
-
- Schnupf P., Portnoy D. A. (2007) Listeriolysin O: a phagosome-specific lysin. Microbes Infect. 9, 1176–1187 - PubMed
-
- Ishino T., Chinzei Y., Yuda M. (2005) A Plasmodium sporozoite protein with a membrane attack complex domain is required for breaching the liver sinusoidal cell layer prior to hepatocyte infection. Cell. Microbiol. 7, 199–208 - PubMed
-
- Schneider M. C., Exley R. M., Ram S., Sim R. B., Tang C. M. (2007) Interactions between Neisseria meningitidis and the complement system. Trends Microbiol. 15, 233–240 - PubMed
-
- Voskoboinik I., Smyth M. J., Trapani J. A. (2006) Perforin-mediated target-cell death and immune homeostasis. Nat. Rev. Immunol. 6, 940–952 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
