

Metalloprotein-based MRI probes
- PMID: 23376346
- PMCID: PMC3716366
- DOI: 10.1016/j.febslet.2013.01.044
Metalloprotein-based MRI probes
Abstract
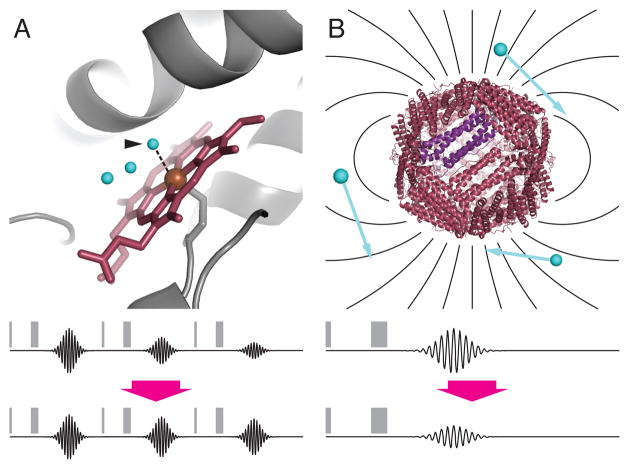
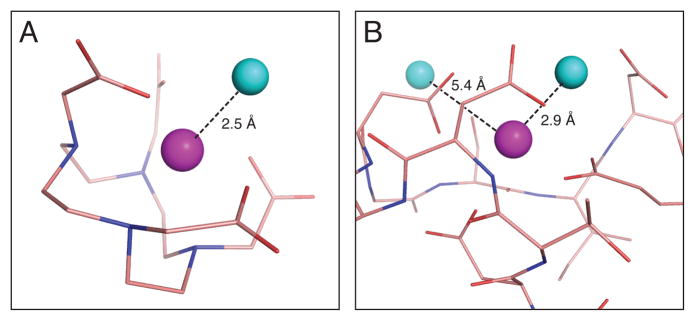
Metalloproteins have long been recognized as key determinants of endogenous contrast in magnetic resonance imaging (MRI) of biological subjects. More recently, both natural and engineered metalloproteins have been harnessed as biotechnological tools to probe gene expression, enzyme activity, and analyte concentrations by MRI. Metalloprotein MRI probes are paramagnetic and function by analogous mechanisms to conventional gadolinium or iron oxide-based MRI contrast agents. Compared with synthetic agents, metalloproteins typically offer worse sensitivity, but the possibilities of using protein engineering and targeted gene expression approaches in conjunction with metalloprotein contrast agents are powerful and sometimes definitive strengths. This review summarizes theoretical and practical aspects of metalloprotein-based contrast agents, and discusses progress in the exploitation of these proteins for molecular imaging applications.
Copyright © 2013 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
Figures


Similar articles
-
Engineering Metalloprotein Functions in Designed and Native Scaffolds.Trends Biochem Sci. 2019 Dec;44(12):1022-1040. doi: 10.1016/j.tibs.2019.06.006. Epub 2019 Jul 13. Trends Biochem Sci. 2019. PMID: 31307903 Review.
-
Magnetite-nanoparticle-encapsulated pH-responsive polymeric micelle as an MRI probe for detecting acidic pathologic areas.Small. 2010 Jun 6;6(11):1201-4. doi: 10.1002/smll.200902317. Small. 2010. PMID: 20449849 No abstract available.
-
Gadolinium-binding helix-turn-helix peptides: DNA-dependent MRI contrast agents.Chem Commun (Camb). 2003 Oct 21;(20):2574-5. doi: 10.1039/b307817e. Chem Commun (Camb). 2003. PMID: 14594287
-
Contrast agents for MRI.Mater Sci Eng C Mater Biol Appl. 2013 Dec 1;33(8):4485-97. doi: 10.1016/j.msec.2013.07.012. Epub 2013 Jul 18. Mater Sci Eng C Mater Biol Appl. 2013. PMID: 24094150 Review.
-
Biomolecular MRI reporters: Evolution of new mechanisms.Prog Nucl Magn Reson Spectrosc. 2017 Nov;102-103:32-42. doi: 10.1016/j.pnmrs.2017.05.002. Epub 2017 Jun 3. Prog Nucl Magn Reson Spectrosc. 2017. PMID: 29157492 Free PMC article. Review.
Cited by
-
Rational design of a genetically encoded NMR zinc sensor.Chem Sci. 2023 Mar 13;14(14):3809-3815. doi: 10.1039/d3sc00437f. eCollection 2023 Apr 5. Chem Sci. 2023. PMID: 37035699 Free PMC article.
-
Non-invasive imaging using reporter genes altering cellular water permeability.Nat Commun. 2016 Dec 23;7:13891. doi: 10.1038/ncomms13891. Nat Commun. 2016. PMID: 28008959 Free PMC article.
-
The design of functional proteins using tensorized energy calculations.Cell Rep Methods. 2023 Aug 15;3(8):100560. doi: 10.1016/j.crmeth.2023.100560. eCollection 2023 Aug 28. Cell Rep Methods. 2023. PMID: 37671023 Free PMC article.
-
Molecular imaging of EGFR/HER2 cancer biomarkers by protein MRI contrast agents.J Biol Inorg Chem. 2014 Feb;19(2):259-70. doi: 10.1007/s00775-013-1076-3. Epub 2013 Dec 24. J Biol Inorg Chem. 2014. PMID: 24366655 Free PMC article. Review.
-
Methods of Measuring Enzyme Activity Ex Vivo and In Vivo.Annu Rev Anal Chem (Palo Alto Calif). 2018 Jun 12;11(1):509-533. doi: 10.1146/annurev-anchem-061417-125619. Epub 2018 Mar 5. Annu Rev Anal Chem (Palo Alto Calif). 2018. PMID: 29505726 Free PMC article. Review.
References
-
- Waldron KJ, Rutherford JC, Ford D, Robinson NJ. Metalloproteins and metal sensing. Nature. 2009;460:823–830. - PubMed
-
- Voet D, Voet JG. Biochemistry. J. Wiley & Sons; New York: 2004.
-
- Hochuli E, Bannwarth W, Döbeli H, Gentz R, Stüber D. Genetic approach to facilitate purification of recombinant proteins with a novel metal chelate adsorbent. Nat Biotechnol. 1988;6:1321–1325.
-
- Hazen TC. Cometabolic Bioremediation. In: Timmis KN, editor. Handbook of Hydrocarbon and Lipid Microbiology. Springer Berlin Heidelberg; Berlin, Heidelberg: 2010. pp. 2505–2514.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
