

In the absence of ATPase activity, pre-RC formation is blocked prior to MCM2-7 hexamer dimerization
- PMID: 23376927
- PMCID: PMC3597701
- DOI: 10.1093/nar/gkt043
In the absence of ATPase activity, pre-RC formation is blocked prior to MCM2-7 hexamer dimerization
Abstract
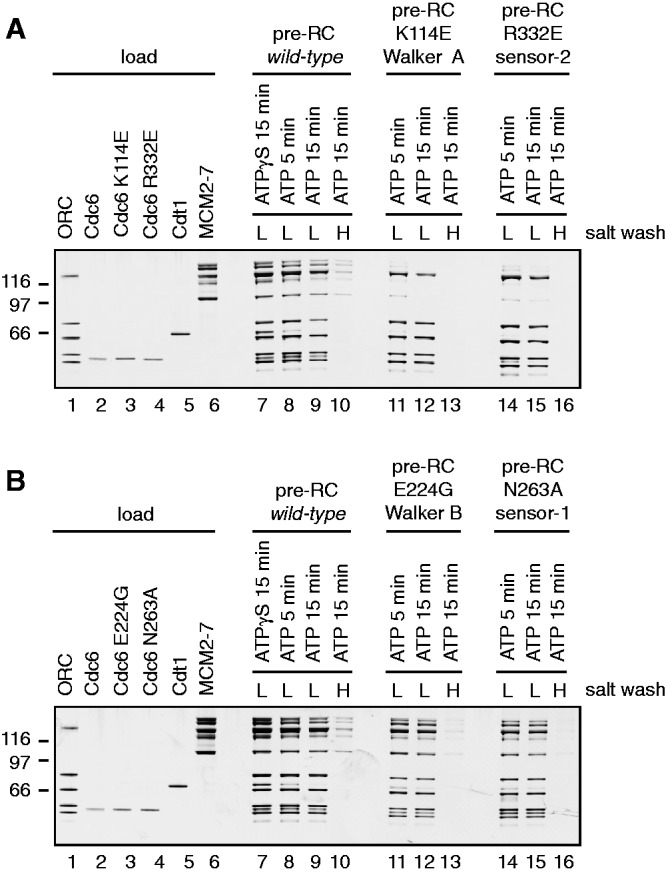
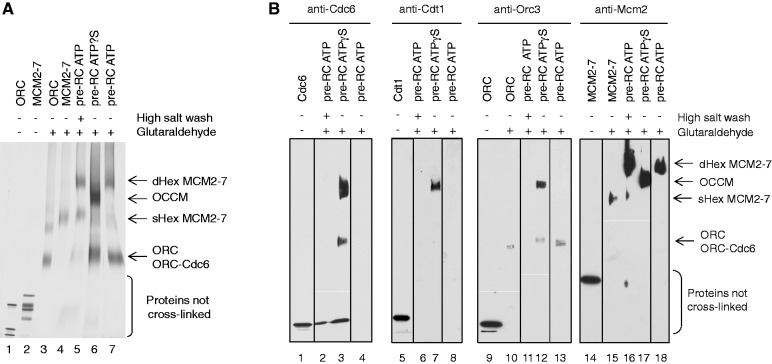
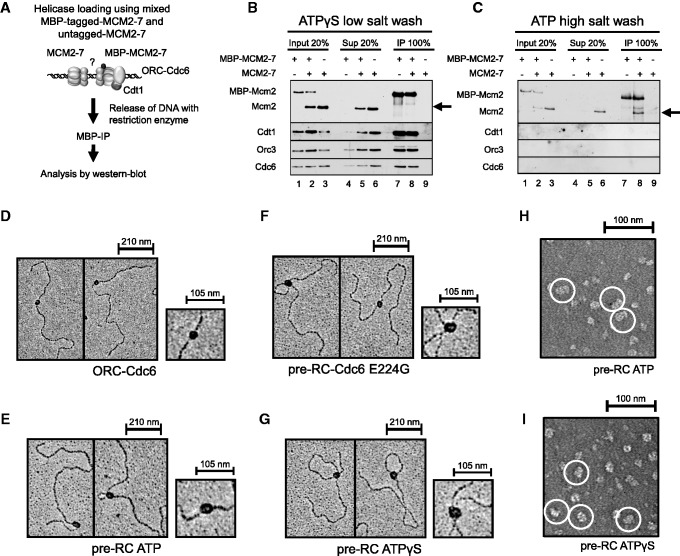
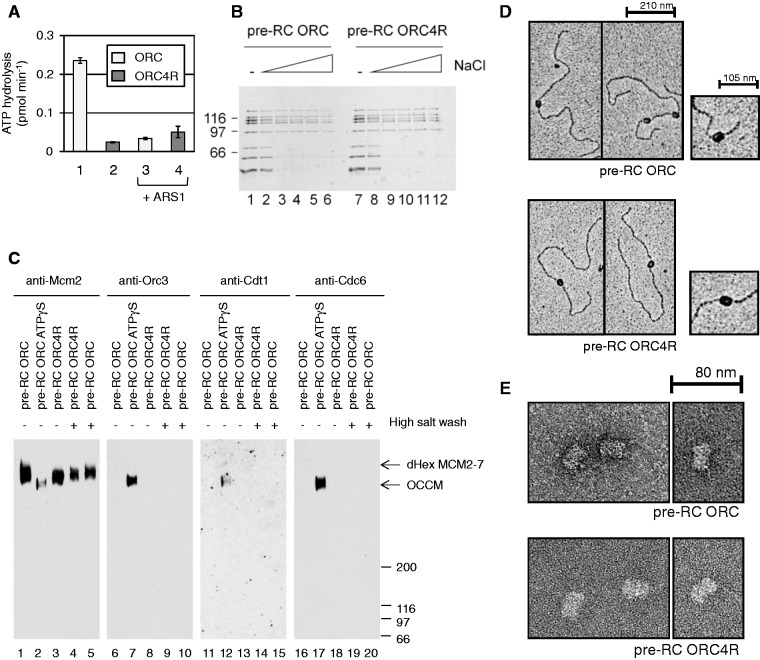
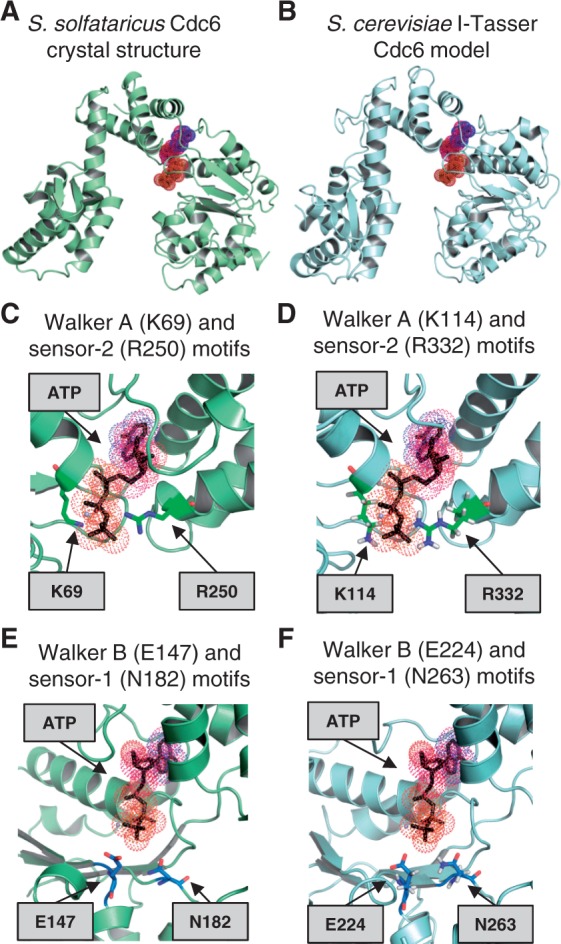
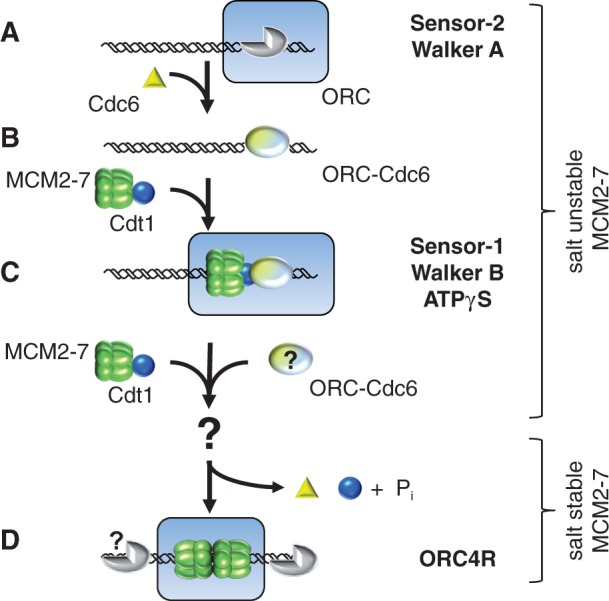
The origin recognition complex (ORC) of Saccharomyces cerevisiae binds origin DNA and cooperates with Cdc6 and Cdt1 to load the replicative helicase MCM2-7 onto DNA. Helicase loading involves two MCM2-7 hexamers that assemble into a double hexamer around double-stranded DNA. This reaction requires ORC and Cdc6 ATPase activity, but it is unknown how these proteins control MCM2-7 double hexamer formation. We demonstrate that mutations in Cdc6 sensor-2 and Walker A motifs, which are predicted to affect ATP binding, influence the ORC-Cdc6 interaction and MCM2-7 recruitment. In contrast, a Cdc6 sensor-1 mutant affects MCM2-7 loading and Cdt1 release, similar as a Cdc6 Walker B ATPase mutant. Moreover, we show that Orc1 ATP hydrolysis is not involved in helicase loading or in releasing ORC from loaded MCM2-7. To determine whether Cdc6 regulates MCM2-7 double hexamer formation, we analysed complex assembly. We discovered that inhibition of Cdc6 ATPase restricts MCM2-7 association with origin DNA to a single hexamer, while active Cdc6 ATPase promotes recruitment of two MCM2-7 hexamer to origin DNA. Our findings illustrate how conserved Cdc6 AAA+ motifs modulate MCM2-7 recruitment, show that ATPase activity is required for MCM2-7 hexamer dimerization and demonstrate that MCM2-7 hexamers are recruited to origins in a consecutive process.
Figures
Similar articles
-
An ORC/Cdc6/MCM2-7 complex is formed in a multistep reaction to serve as a platform for MCM double-hexamer assembly.Mol Cell. 2013 May 23;50(4):577-88. doi: 10.1016/j.molcel.2013.03.026. Epub 2013 Apr 18. Mol Cell. 2013. PMID: 23603117
-
The ORC/Cdc6/MCM2-7 complex facilitates MCM2-7 dimerization during prereplicative complex formation.Nucleic Acids Res. 2014 Feb;42(4):2257-69. doi: 10.1093/nar/gkt1148. Epub 2013 Nov 14. Nucleic Acids Res. 2014. PMID: 24234446 Free PMC article.
-
Cryo-EM structure of a helicase loading intermediate containing ORC-Cdc6-Cdt1-MCM2-7 bound to DNA.Nat Struct Mol Biol. 2013 Aug;20(8):944-51. doi: 10.1038/nsmb.2629. Epub 2013 Jul 14. Nat Struct Mol Biol. 2013. PMID: 23851460 Free PMC article.
-
DnaA, ORC, and Cdc6: similarity beyond the domains of life and diversity.Biochem Cell Biol. 2010 Feb;88(1):49-62. doi: 10.1139/o09-154. Biochem Cell Biol. 2010. PMID: 20130679 Review.
-
Unique Roles of the Non-identical MCM Subunits in DNA Replication Licensing.Mol Cell. 2017 Jul 20;67(2):168-179. doi: 10.1016/j.molcel.2017.06.016. Mol Cell. 2017. PMID: 28732205 Review.
Cited by
-
A new class of disordered elements controls DNA replication through initiator self-assembly.Elife. 2019 Sep 27;8:e48562. doi: 10.7554/eLife.48562. Elife. 2019. PMID: 31560342 Free PMC article.
-
Switch on the engine: how the eukaryotic replicative helicase MCM2-7 becomes activated.Chromosoma. 2015 Mar;124(1):13-26. doi: 10.1007/s00412-014-0489-2. Epub 2014 Oct 12. Chromosoma. 2015. PMID: 25308420 Review.
-
Structural basis of Mcm2-7 replicative helicase loading by ORC-Cdc6 and Cdt1.Nat Struct Mol Biol. 2017 Mar;24(3):316-324. doi: 10.1038/nsmb.3372. Epub 2017 Feb 13. Nat Struct Mol Biol. 2017. PMID: 28191893 Free PMC article.
-
The High-Affinity Interaction between ORC and DNA that Is Required for Replication Licensing Is Inhibited by 2-Arylquinolin-4-Amines.Cell Chem Biol. 2017 Aug 17;24(8):981-992.e4. doi: 10.1016/j.chembiol.2017.06.019. Epub 2017 Aug 3. Cell Chem Biol. 2017. PMID: 28781123 Free PMC article.
-
Mechanisms and regulation of DNA replication initiation in eukaryotes.Crit Rev Biochem Mol Biol. 2017 Apr;52(2):107-144. doi: 10.1080/10409238.2016.1274717. Epub 2017 Jan 17. Crit Rev Biochem Mol Biol. 2017. PMID: 28094588 Free PMC article. Review.
References
-
- Bell SP, Stillman B. ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex. Nature. 1992;357:128–134. - PubMed
-
- Remus D, Diffley JF. Eukaryotic DNA replication control: lock and load, then fire. Curr. Opin. Cell Biol. 2009;21:771–777. - PubMed
-
- Randell JC, Bowers JL, Rodriguez HK, Bell SP. Sequential ATP hydrolysis by Cdc6 and ORC directs loading of the MCM2–7 helicase. Mol. Cell. 2006;21:29–39. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
