

Dicer-independent processing of short hairpin RNAs
- PMID: 23376931
- PMCID: PMC3616727
- DOI: 10.1093/nar/gkt036
Dicer-independent processing of short hairpin RNAs
Abstract
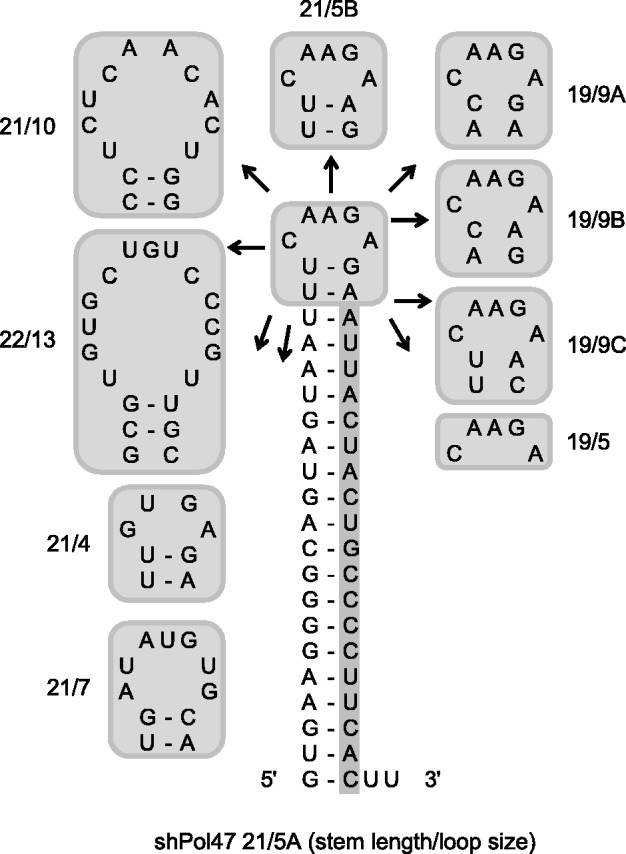
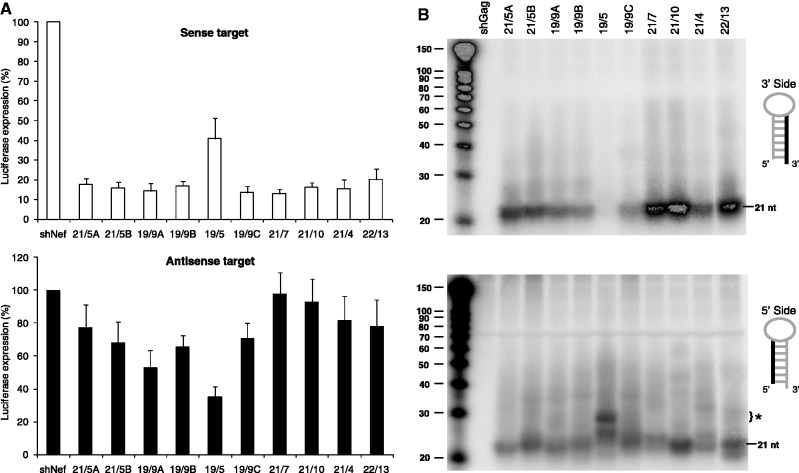
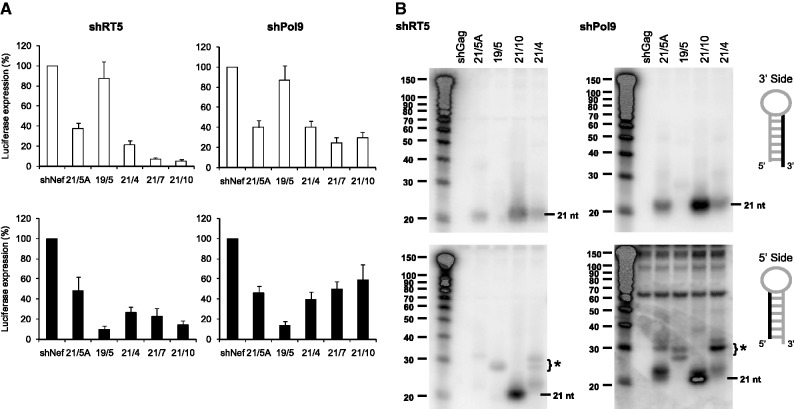
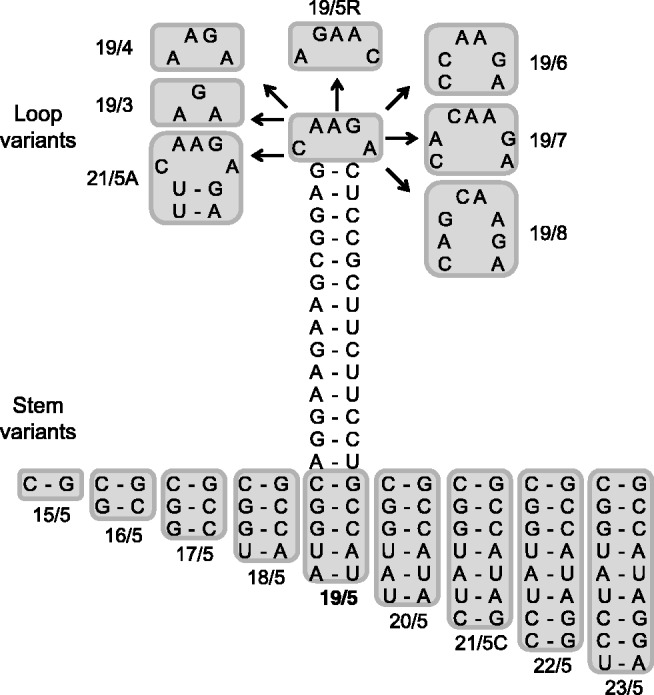
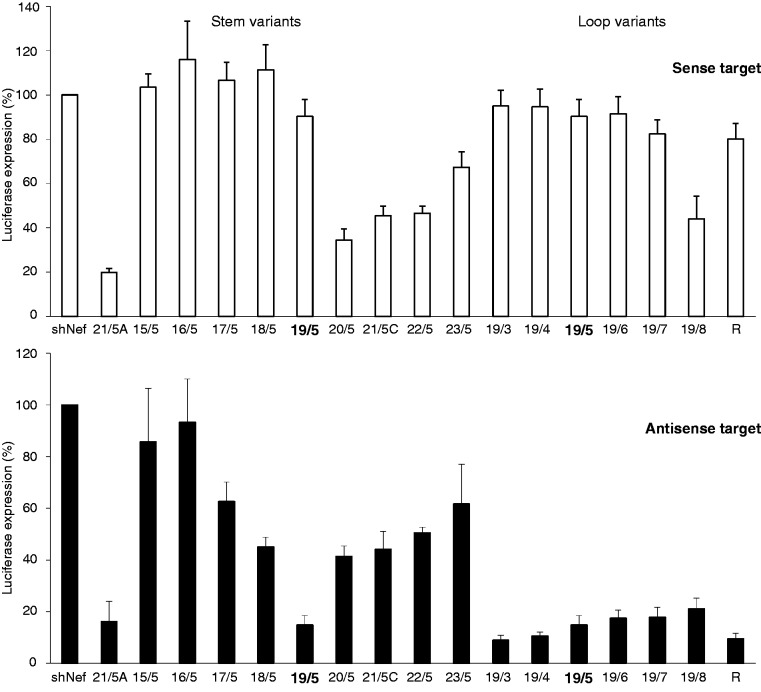
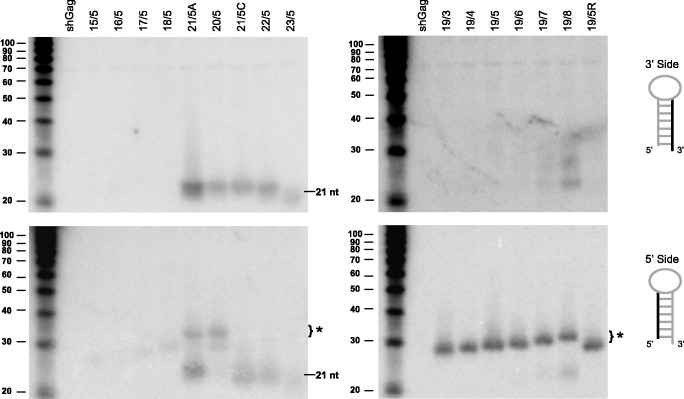
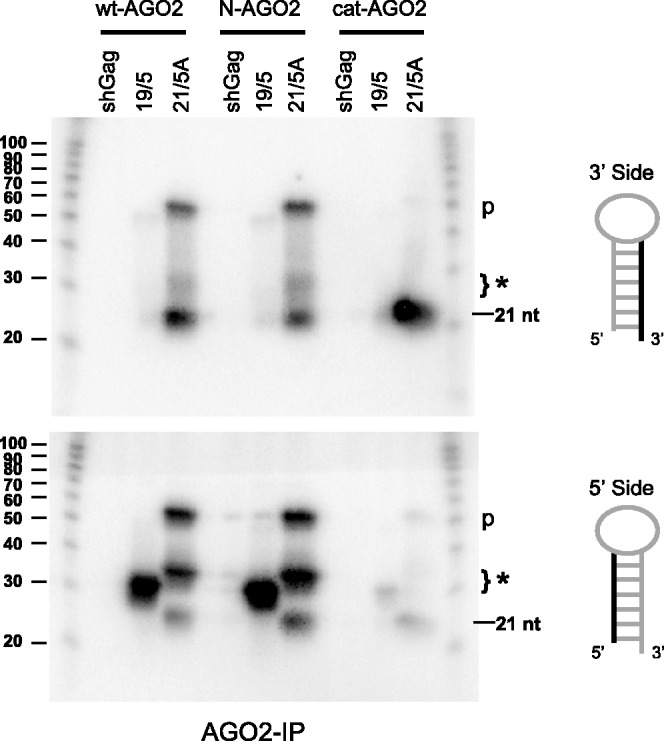
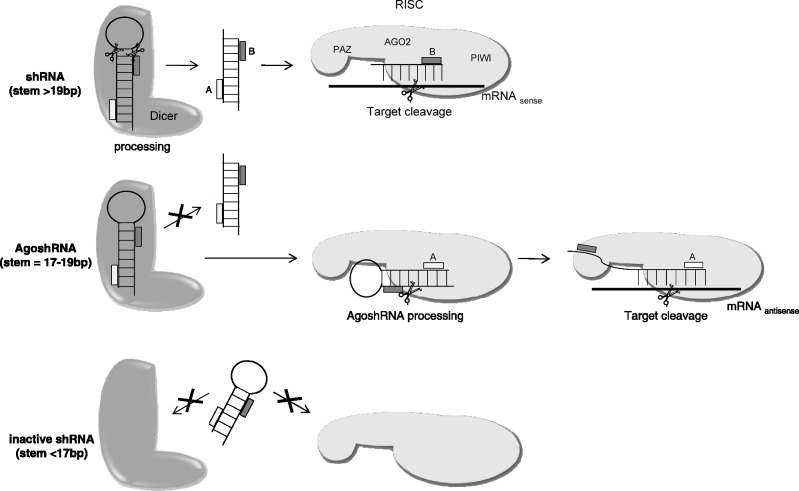
Short hairpin RNAs (shRNAs) are widely used to induce RNA interference (RNAi). We tested a variety of shRNAs that differed in stem length and terminal loop size and revealed strikingly different RNAi activities and shRNA-processing patterns. Interestingly, we identified a specific shRNA design that uses an alternative Dicer-independent processing pathway. Detailed analyses indicated that a short shRNA stem length is critical for avoiding Dicer processing and activation of the alternative processing route, in which the shRNA is incorporated into RISC and processed by the AGO2-mediated slicer activity. Such alternatively processed shRNAs (AgoshRNAs) yield only a single RNA strand that effectively induces RNAi, whereas conventional shRNA processing results in an siRNA duplex of which both strands can trigger RNAi. Both the processing and subsequent RNAi activity of these AgoshRNAs are thus mediated by the RISC-component AGO2. These results have important implications for the future design of more specific RNAi therapeutics.
Figures
References
-
- Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 1998;391:806–811. - PubMed
-
- Bernstein E, Caudy AA, Hammond SM, Hannon GJ. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 2001;409:363–366. - PubMed
-
- Nykanen A, Haley B, Zamore PD. ATP requirements and small interfering RNA structure in the RNA interference pathway. Cell. 2001;107:309–321. - PubMed
-
- Denli AM, Tops BB, Plasterk RH, Ketting RF, Hannon GJ. Processing of primary microRNAs by the microprocessor complex. Nature. 2004;432:231–235. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
