

The Tec kinase ITK regulates thymic expansion, emigration, and maturation of γδ NKT cells
- PMID: 23378428
- PMCID: PMC3594397
- DOI: 10.4049/jimmunol.1202531
The Tec kinase ITK regulates thymic expansion, emigration, and maturation of γδ NKT cells
Abstract
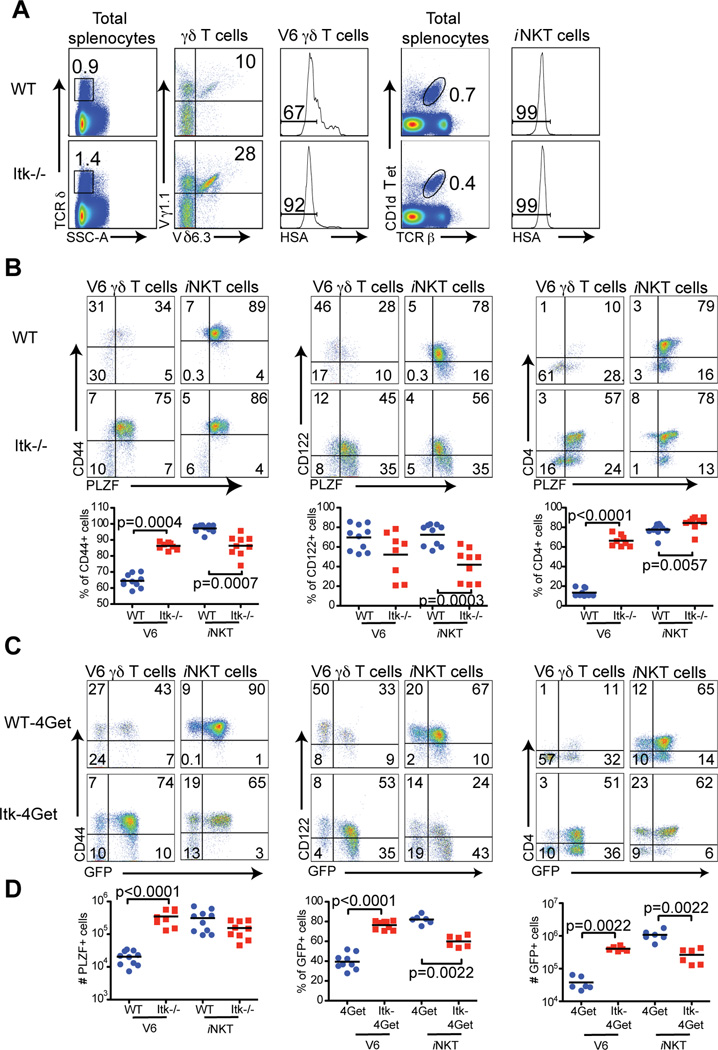
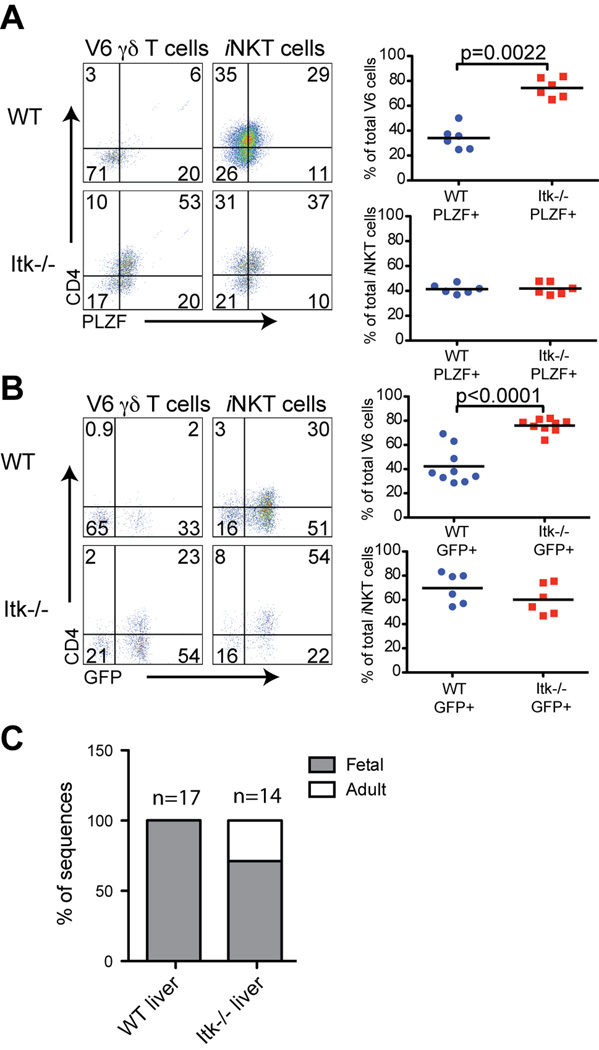
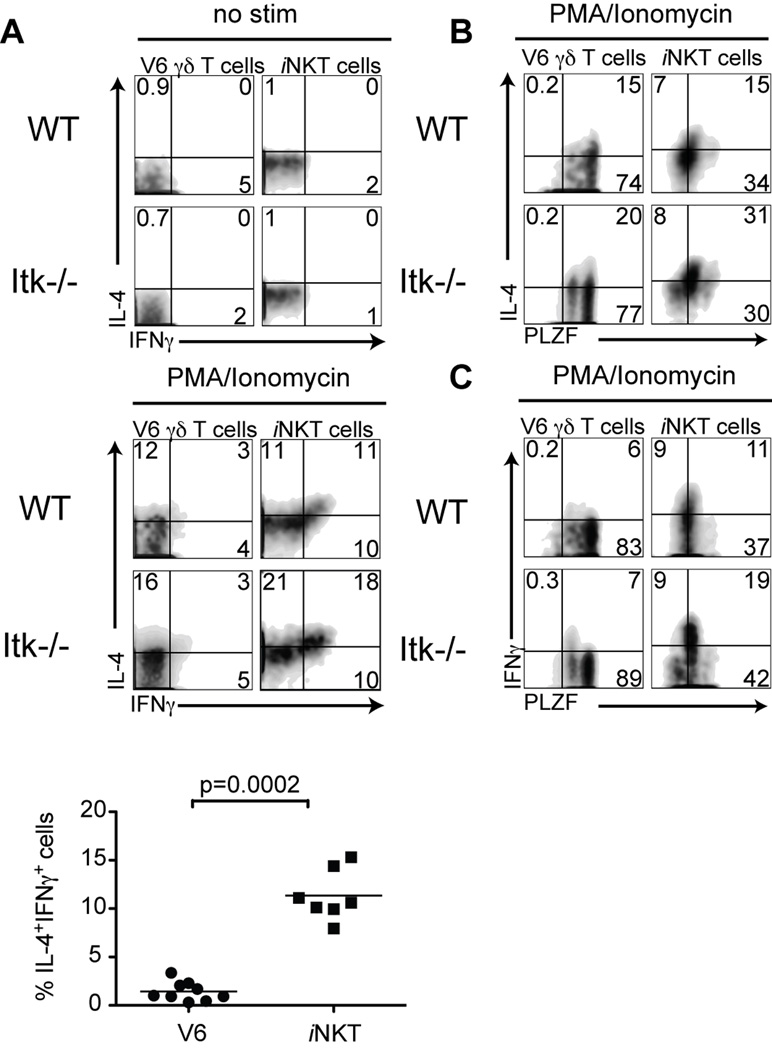
The Tec family tyrosine kinase, Itk, regulates signaling downstream of the TCR. The absence of Itk in CD4(+) T cells results in impaired Th2 responses along with defects in maturation, cytokine production, and survival of iNKT cells. Paradoxically, Itk(-/-) mice have spontaneously elevated serum IgE levels, resulting from an expansion of the Vγ1.1(+)Vδ6.3(+) subset of γδ T cells, known as γδ NKT cells. Comparisons between γδ NKT cells and αβ iNKT cells showed convergence in the pattern of cell surface marker expression, cytokine profiles, and gene expression, suggesting that these two subsets of NKT cells undergo similar differentiation programs. Hepatic γδ NKT cells have an invariant TCR and are derived predominantly from fetal progenitors that expand in the thymus during the first weeks of life. The adult thymus contains these invariant γδ NKT cells plus a heterogeneous population of Vγ1.1(+)Vδ6.3(+) T cells with diverse CDR3 sequences. This latter population, normally excluded from the liver, escapes the thymus and homes to the liver when Itk is absent. In addition, Itk(-/-) γδ NKT cells persistently express high levels of Zbtb16 (PLZF) and Il4, genes that are normally downregulated in the most mature subsets of NKT cells. These data indicate that Itk signaling is required to prevent the expansion of γδ NKT cells in the adult thymus, to block their emigration, and to promote terminal NKT cell maturation.
Figures





Similar articles
-
Tec kinase Itk in gammadeltaT cells is pivotal for controlling IgE production in vivo.Proc Natl Acad Sci U S A. 2009 May 19;106(20):8308-13. doi: 10.1073/pnas.0808459106. Epub 2009 May 1. Proc Natl Acad Sci U S A. 2009. PMID: 19416854 Free PMC article.
-
Enhanced development of CD4+ gammadelta T cells in the absence of Itk results in elevated IgE production.Blood. 2009 Jul 16;114(3):564-71. doi: 10.1182/blood-2008-12-196345. Epub 2009 May 14. Blood. 2009. PMID: 19443662 Free PMC article.
-
MicroRNA-181a/b-1 Is Not Required for Innate γδ NKT Effector Cell Development.PLoS One. 2015 Dec 16;10(12):e0145010. doi: 10.1371/journal.pone.0145010. eCollection 2015. PLoS One. 2015. PMID: 26673421 Free PMC article.
-
Tec family kinases: Itk signaling and the development of NKT αβ and γδ T cells.FEBS J. 2011 Jun;278(12):1970-9. doi: 10.1111/j.1742-4658.2011.08074.x. Epub 2011 Mar 29. FEBS J. 2011. PMID: 21362141 Review.
-
Thymus medulla fosters generation of natural Treg cells, invariant γδ T cells, and invariant NKT cells: what we learn from intrathymic migration.Eur J Immunol. 2015 Mar;45(3):652-60. doi: 10.1002/eji.201445108. Epub 2015 Feb 13. Eur J Immunol. 2015. PMID: 25615828 Free PMC article. Review.
Cited by
-
Interleukin-2-Inducible T-Cell Kinase Deficiency Impairs Early Pulmonary Protection Against Mycobacterium tuberculosis Infection.Front Immunol. 2020 Jan 24;10:3103. doi: 10.3389/fimmu.2019.03103. eCollection 2019. Front Immunol. 2020. PMID: 32038633 Free PMC article.
-
Development of innate CD4+ and CD8+ T cells in Itk-deficient mice is regulated by distinct pathways.J Immunol. 2014 Jul 15;193(2):688-99. doi: 10.4049/jimmunol.1302059. Epub 2014 Jun 18. J Immunol. 2014. PMID: 24943215 Free PMC article.
-
The Potent ITK/BTK Inhibitor Ibrutinib Is Effective for the Treatment of Experimental Visceral Leishmaniasis Caused by Leishmania donovani.J Infect Dis. 2019 Jan 29;219(4):599-608. doi: 10.1093/infdis/jiy552. J Infect Dis. 2019. PMID: 30239895 Free PMC article.
-
IL-2-inducible T cell kinase tunes T regulatory cell development and is required for suppressive function.J Immunol. 2014 Sep 1;193(5):2267-72. doi: 10.4049/jimmunol.1400968. Epub 2014 Jul 25. J Immunol. 2014. PMID: 25063868 Free PMC article.
-
Innate PLZF+CD4+ αβ T cells develop and expand in the absence of Itk.J Immunol. 2014 Jul 15;193(2):673-87. doi: 10.4049/jimmunol.1302058. Epub 2014 Jun 13. J Immunol. 2014. PMID: 24928994 Free PMC article.
References
-
- Kaech SM, Wherry EJ, Ahmed R. Effector and memory T-cell differentiation: implications for vaccine development. Nat. Rev. Immunol. 2002;2:251–262. - PubMed
-
- Surh CD, Sprent J. Regulation of mature T cell homeostasis. Seminars in Immunology. 2005;17:183–191. - PubMed
-
- Jameson SC. T cell homeostasis: keeping useful T cells alive and live T cells useful. Seminars in Immunology. 2005;17:231–237. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
