

Root-root interactions: extending our perspective to be more inclusive of the range of theories in ecology and agriculture using in-vivo analyses
- PMID: 23378521
- PMCID: PMC3698385
- DOI: 10.1093/aob/mcs296
Root-root interactions: extending our perspective to be more inclusive of the range of theories in ecology and agriculture using in-vivo analyses
Abstract
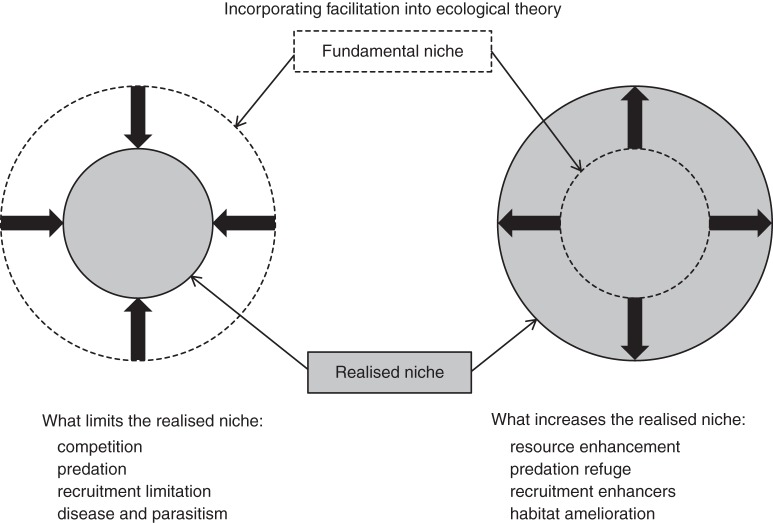
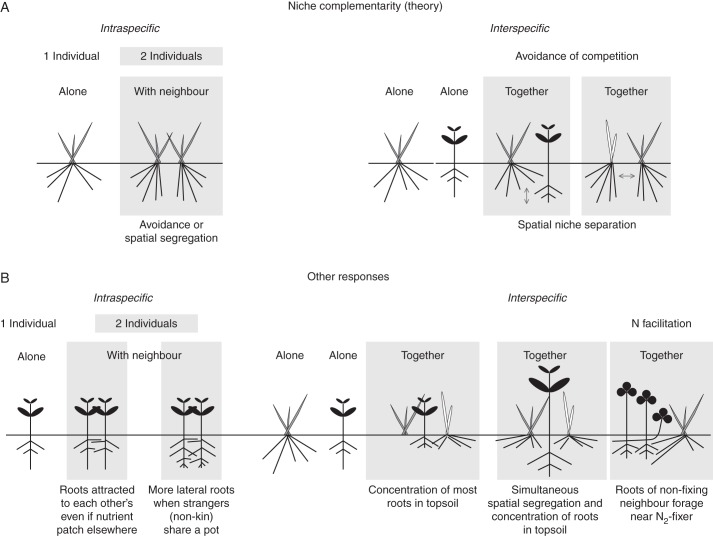
Background: There is a large body of literature on competitive interactions among plants, but many studies have only focused on above-ground interactions and little is known about root-root dynamics between interacting plants. The perspective on possible mechanisms that explain the outcome of root-root interactions has recently been extended to include non-resource-driven mechanisms (as well as resource-driven mechanisms) of root competition and positive interactions such as facilitation. These approaches have often suffered from being static, partly due to the lack of appropriate methodologies for in-situ non-destructive root characterization.
Scope: Recent studies show that interactive effects of plant neighbourhood interactions follow non-linear and non-additive paths that are hard to explain. Common outcomes such as accumulation of roots mainly in the topsoil cannot be explained solely by competition theory but require a more inclusive theoretical, as well as an improved methodological framework. This will include the question of whether we can apply the same conceptual framework to crop versus natural species.
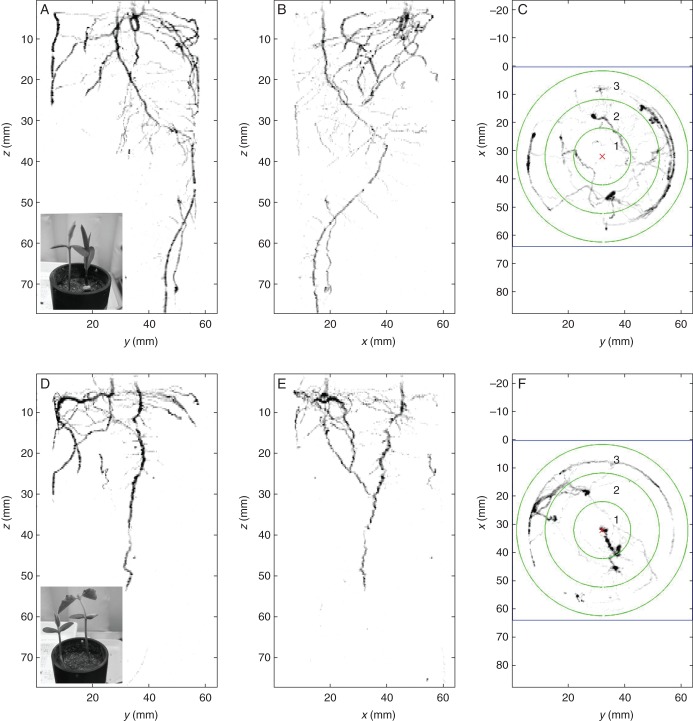
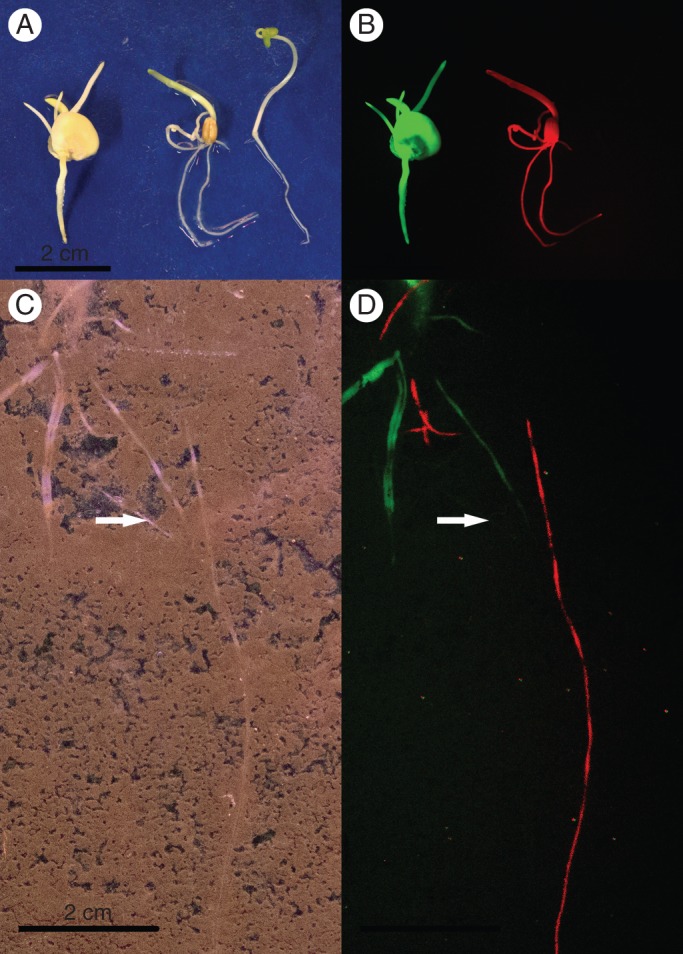
Conclusions: The development of non-invasive methods to dynamically study root-root interactions in vivo will provide the necessary tools to study a more inclusive conceptual framework for root-root interactions. By following the dynamics of root-root interactions through time in a whole range of scenarios and systems, using a wide variety of non-invasive methods, (such as fluorescent protein which now allows us to separately identify the roots of several individuals within soil), we will be much better equipped to answer some of the key questions in root physiology, ecology and agronomy.
Keywords: Root; competition; facilitation; green fluorescent protein; interaction; interspecific; intraspecific; nuclear magnetic resonance; positron emission tomography; red fluorescent protein; resource; rhizotrons.
Figures

Similar articles
-
Rapid transgenerational adaptation in response to intercropping reduces competition.Elife. 2022 Sep 13;11:e77577. doi: 10.7554/eLife.77577. Elife. 2022. PMID: 36097813 Free PMC article.
-
Future paths for the 'exploitative segregation of plant roots' model.Plant Signal Behav. 2021 May 4;16(5):1891755. doi: 10.1080/15592324.2021.1891755. Epub 2021 Mar 1. Plant Signal Behav. 2021. PMID: 33641625 Free PMC article.
-
Plant intraspecific competition and growth stage alter carbon and nitrogen mineralization in the rhizosphere.Plant Cell Environ. 2021 Apr;44(4):1231-1242. doi: 10.1111/pce.13945. Epub 2020 Dec 15. Plant Cell Environ. 2021. PMID: 33175402
-
Detect thy family: Mechanisms, ecology and agricultural aspects of kin recognition in plants.Plant Cell Environ. 2021 Apr;44(4):1059-1071. doi: 10.1111/pce.14011. Epub 2021 Feb 18. Plant Cell Environ. 2021. PMID: 33522615 Free PMC article. Review.
-
Root exudate signals in plant-plant interactions.Plant Cell Environ. 2021 Apr;44(4):1044-1058. doi: 10.1111/pce.13892. Epub 2020 Oct 7. Plant Cell Environ. 2021. PMID: 32931018 Review.
Cited by
-
Belowground neighbor perception in Arabidopsis thaliana studied by transcriptome analysis: roots of Hieracium pilosella cause biotic stress.Front Plant Sci. 2013 Aug 13;4:296. doi: 10.3389/fpls.2013.00296. eCollection 2013. Front Plant Sci. 2013. PMID: 23967000 Free PMC article.
-
Electrical Capacitance versus Minirhizotron Technique: A Study of Root Dynamics in Wheat-Pea Intercrops.Plants (Basel). 2021 Sep 23;10(10):1991. doi: 10.3390/plants10101991. Plants (Basel). 2021. PMID: 34685800 Free PMC article.
-
Arguments for and against self and non-self root recognition in plants.Front Plant Sci. 2014 Nov 6;5:614. doi: 10.3389/fpls.2014.00614. eCollection 2014. Front Plant Sci. 2014. PMID: 25414719 Free PMC article.
-
Auxin and the integration of environmental signals into plant root development.Ann Bot. 2013 Dec;112(9):1655-65. doi: 10.1093/aob/mct229. Epub 2013 Oct 17. Ann Bot. 2013. PMID: 24136877 Free PMC article. Review.
-
Extracting multiple interacting root systems using X-ray microcomputed tomography.Plant J. 2015 Dec;84(5):1034-43. doi: 10.1111/tpj.13047. Plant J. 2015. PMID: 26461469 Free PMC article.
References
-
- Ashton IW, Miller AE, Bowman WD, Suding KN. Niche complementarity due to plasticity in resource use: plant partitioning of chemical N forms. Ecology. 2010;91:3252–3260. - PubMed
-
- Bessler H, Temperton VM, Roscher C, et al. Aboveground overyielding in grassland mixtures is associated with reduced biomass partitioning to belowground organs. Ecology. 2009;90:1520–1530. - PubMed
-
- Bezemer TM, Fountain MT, Barea JM, et al. Divergent composition but similar function of soil food webs of individual plants: plant species and community effects. Ecology. 2010;91:3027–3036. - PubMed
-
- Brooker R, Kikvidze Z, Pugnaire FI, et al. The importance of importance. Oikos. 2005;109:63–70.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
