

Intrinsic evolutionary constraints on protease structure, enzyme acylation, and the identity of the catalytic triad
- PMID: 23382230
- PMCID: PMC3581919
- DOI: 10.1073/pnas.1221050110
Intrinsic evolutionary constraints on protease structure, enzyme acylation, and the identity of the catalytic triad
Abstract
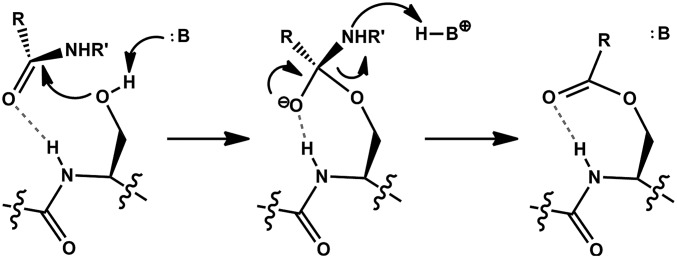
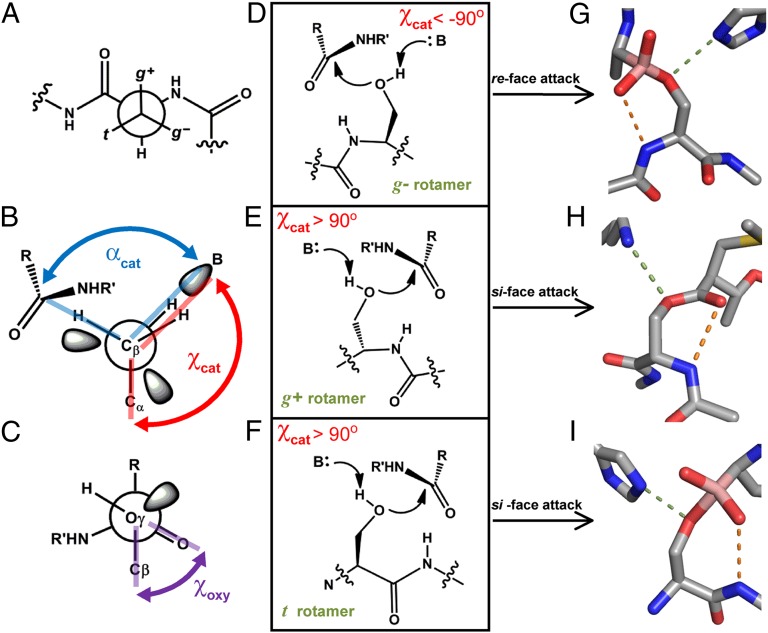
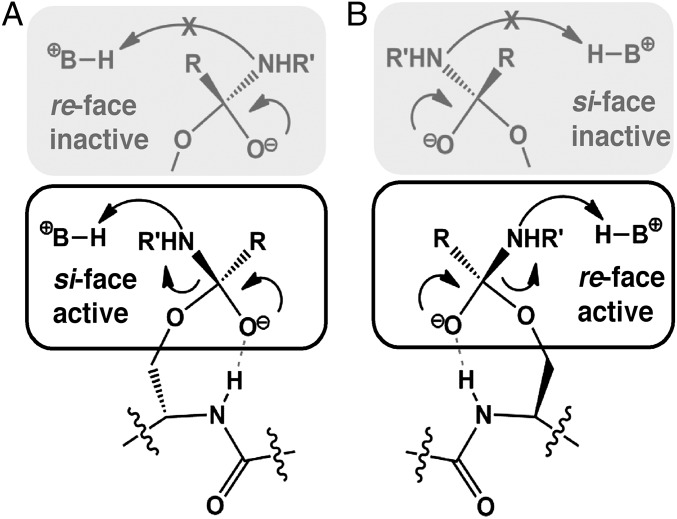
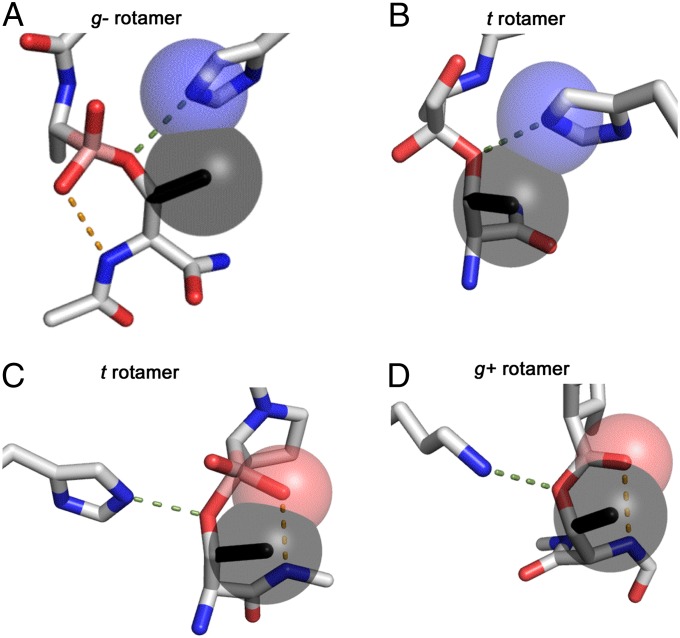
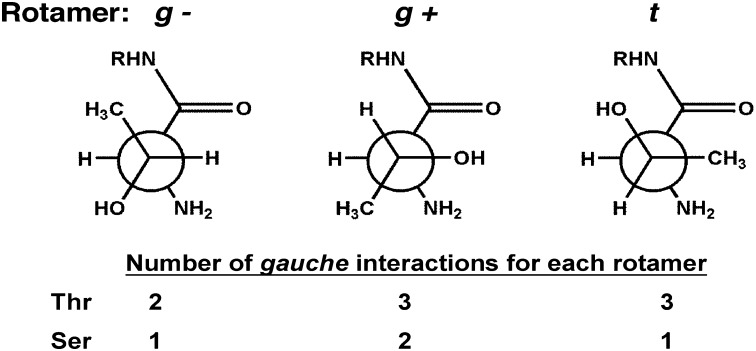
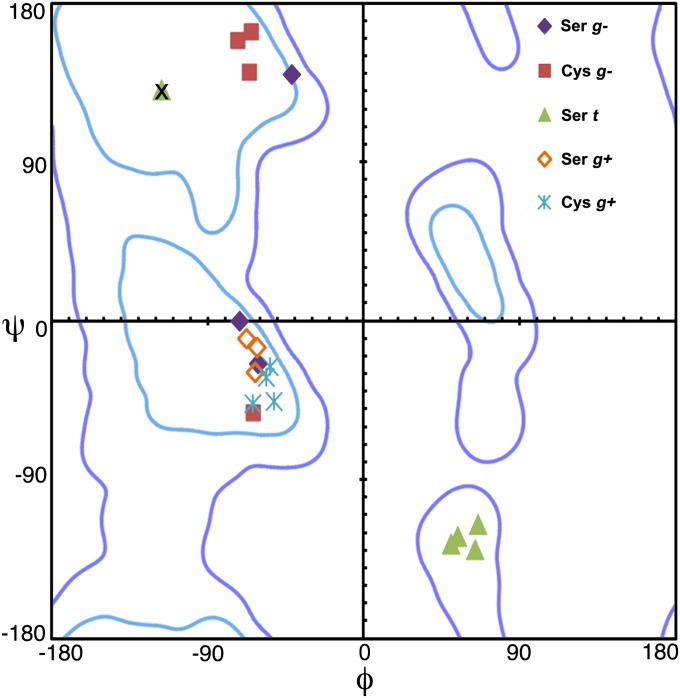
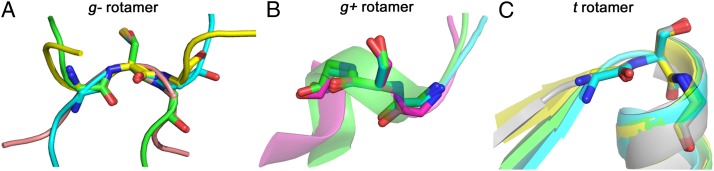
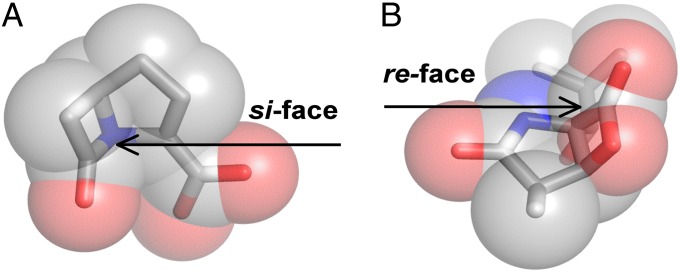
The study of proteolysis lies at the heart of our understanding of biocatalysis, enzyme evolution, and drug development. To understand the degree of natural variation in protease active sites, we systematically evaluated simple active site features from all serine, cysteine and threonine proteases of independent lineage. This convergent evolutionary analysis revealed several interrelated and previously unrecognized relationships. The reactive rotamer of the nucleophile determines which neighboring amide can be used in the local oxyanion hole. Each rotamer-oxyanion hole combination limits the location of the moiety facilitating proton transfer and, combined together, fixes the stereochemistry of catalysis. All proteases that use an acyl-enzyme mechanism naturally divide into two classes according to which face of the peptide substrate is attacked during catalysis. We show that each class is subject to unique structural constraints that have governed the convergent evolution of enzyme structure. Using this framework, we show that the γ-methyl of Thr causes an intrinsic steric clash that precludes its use as the nucleophile in the traditional catalytic triad. This constraint is released upon autoproteolysis and we propose a molecular basis for the increased enzymatic efficiency introduced by the γ-methyl of Thr. Finally, we identify several classes of natural products whose mode of action is sensitive to the division according to the face of attack identified here. This analysis of protease structure and function unifies 50 y of biocatalysis research, providing a framework for the continued study of enzyme evolution and the development of inhibitors with increased selectivity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Warshel A, Naray-Szabo G, Sussman F, Hwang JK. How do serine proteases really work? Biochemistry. 1989;28(9):3629–3637. - PubMed
-
- Fersht A. Structure and Mechanism in Protein Science: A Guide to Enzyme Catalysis and Protein Folding. New York: Freeman; 1999.
-
- Kraut J. How do enzymes work? Science. 1988;242(4878):533–540. - PubMed
-
- Warshel A. Electrostatic origin of the catalytic power of enzymes and the role of preorganized active sites. J Biol Chem. 1998;273(42):27035–27038. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
