

Bayesian inference of spatial organizations of chromosomes
- PMID: 23382666
- PMCID: PMC3561073
- DOI: 10.1371/journal.pcbi.1002893
Bayesian inference of spatial organizations of chromosomes
Abstract
Knowledge of spatial chromosomal organizations is critical for the study of transcriptional regulation and other nuclear processes in the cell. Recently, chromosome conformation capture (3C) based technologies, such as Hi-C and TCC, have been developed to provide a genome-wide, three-dimensional (3D) view of chromatin organization. Appropriate methods for analyzing these data and fully characterizing the 3D chromosomal structure and its structural variations are still under development. Here we describe a novel Bayesian probabilistic approach, denoted as "Bayesian 3D constructor for Hi-C data" (BACH), to infer the consensus 3D chromosomal structure. In addition, we describe a variant algorithm BACH-MIX to study the structural variations of chromatin in a cell population. Applying BACH and BACH-MIX to a high resolution Hi-C dataset generated from mouse embryonic stem cells, we found that most local genomic regions exhibit homogeneous 3D chromosomal structures. We further constructed a model for the spatial arrangement of chromatin, which reveals structural properties associated with euchromatic and heterochromatic regions in the genome. We observed strong associations between structural properties and several genomic and epigenetic features of the chromosome. Using BACH-MIX, we further found that the structural variations of chromatin are correlated with these genomic and epigenetic features. Our results demonstrate that BACH and BACH-MIX have the potential to provide new insights into the chromosomal architecture of mammalian cells.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
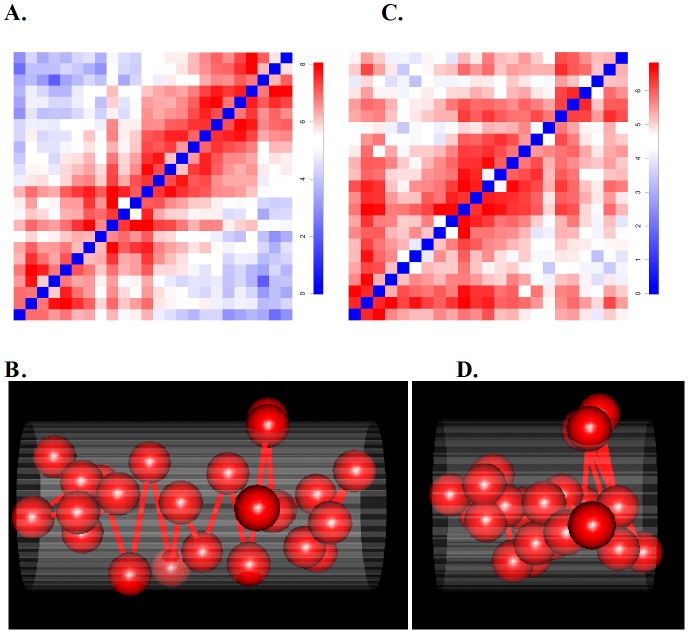
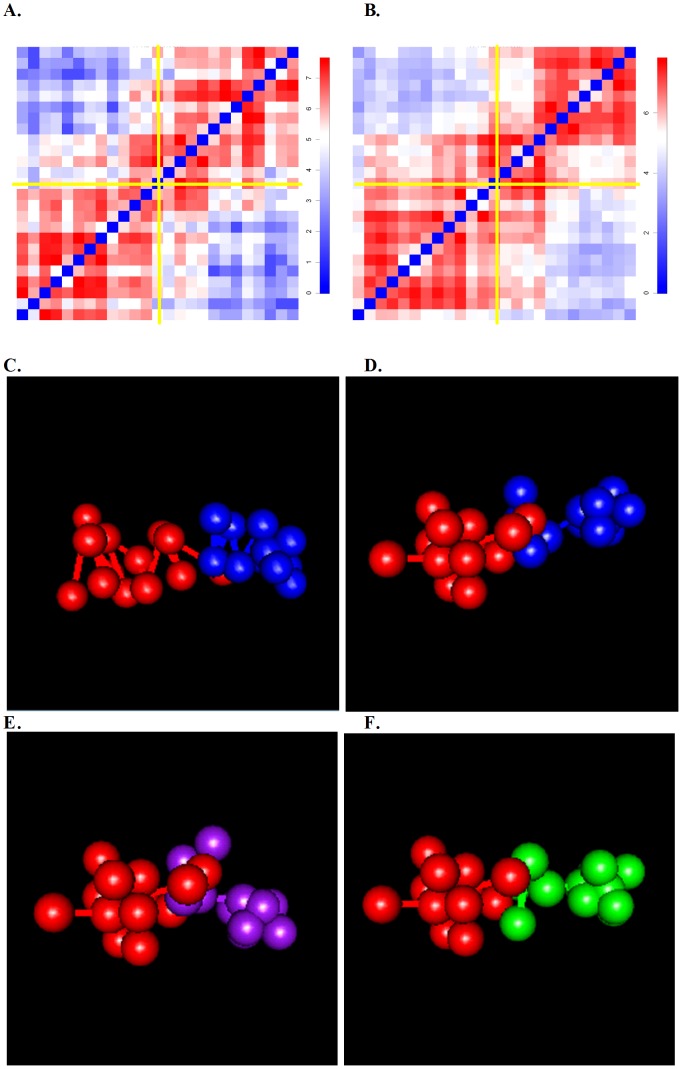

References
-
- Fraser P, Bickmore W (2007) Nuclear organization of the genome and the potential for gene regulation. Nature 447: 413–417. - PubMed
-
- Misteli T (2007) Beyond the sequence: cellular organization of genome function. Cell 128: 787–800. - PubMed
-
- Misteli T (2004) Spatial positioning; a new dimension in genome function. Cell 119: 153–156. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
