

Spiegelzymes: sequence specific hydrolysis of L-RNA with mirror image hammerhead ribozymes and DNAzymes
- PMID: 23382952
- PMCID: PMC3559883
- DOI: 10.1371/journal.pone.0054741
Spiegelzymes: sequence specific hydrolysis of L-RNA with mirror image hammerhead ribozymes and DNAzymes
Abstract
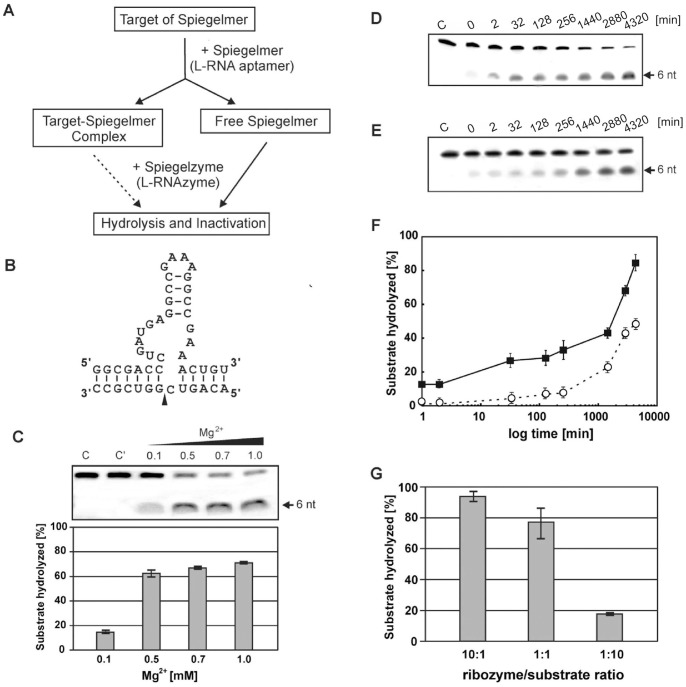
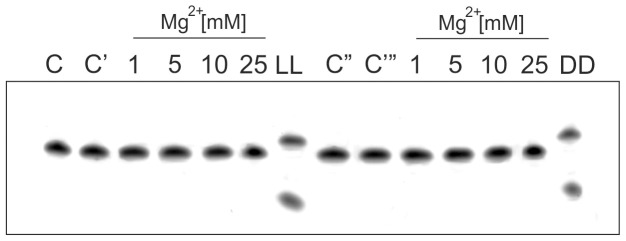
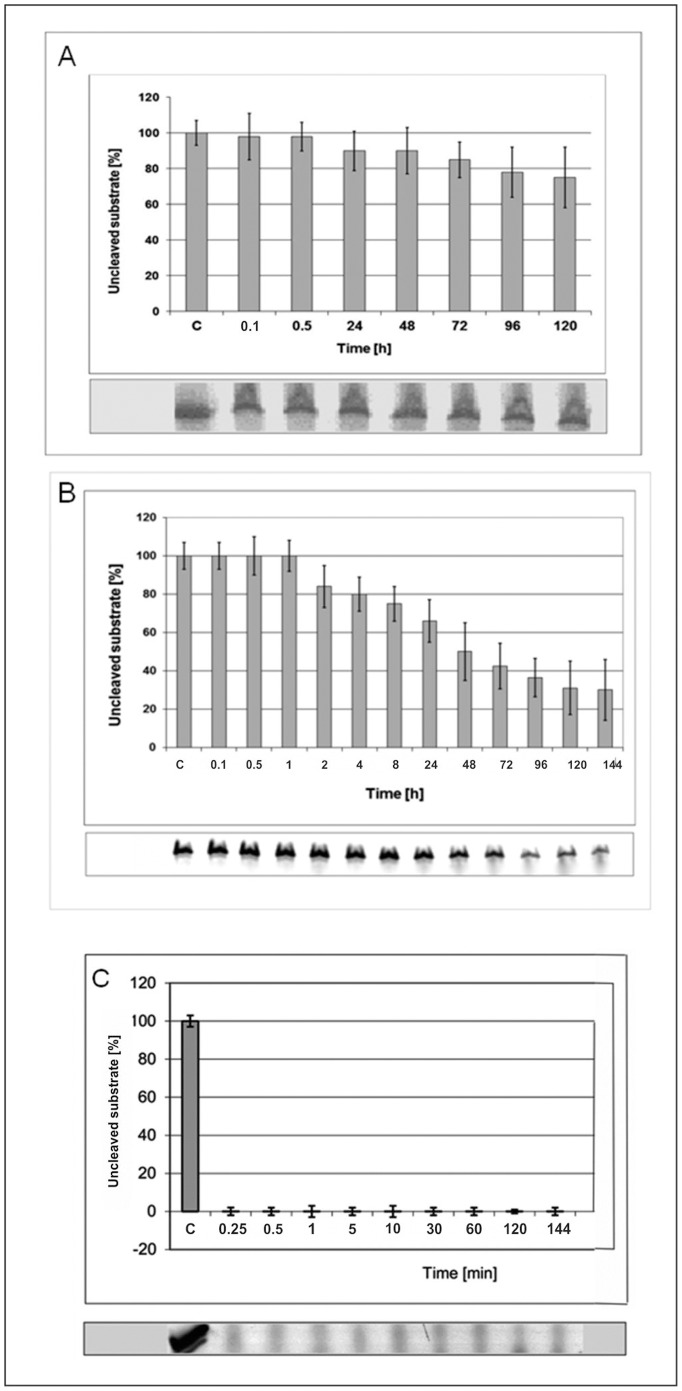
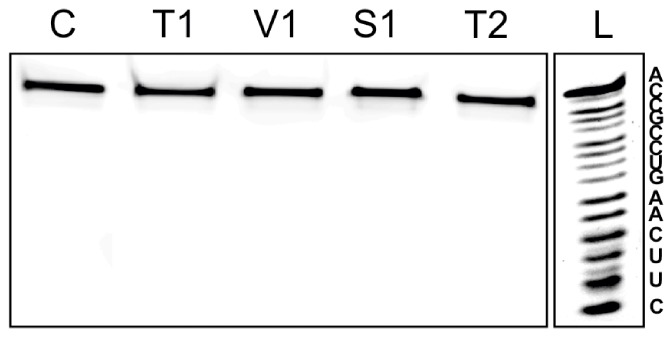
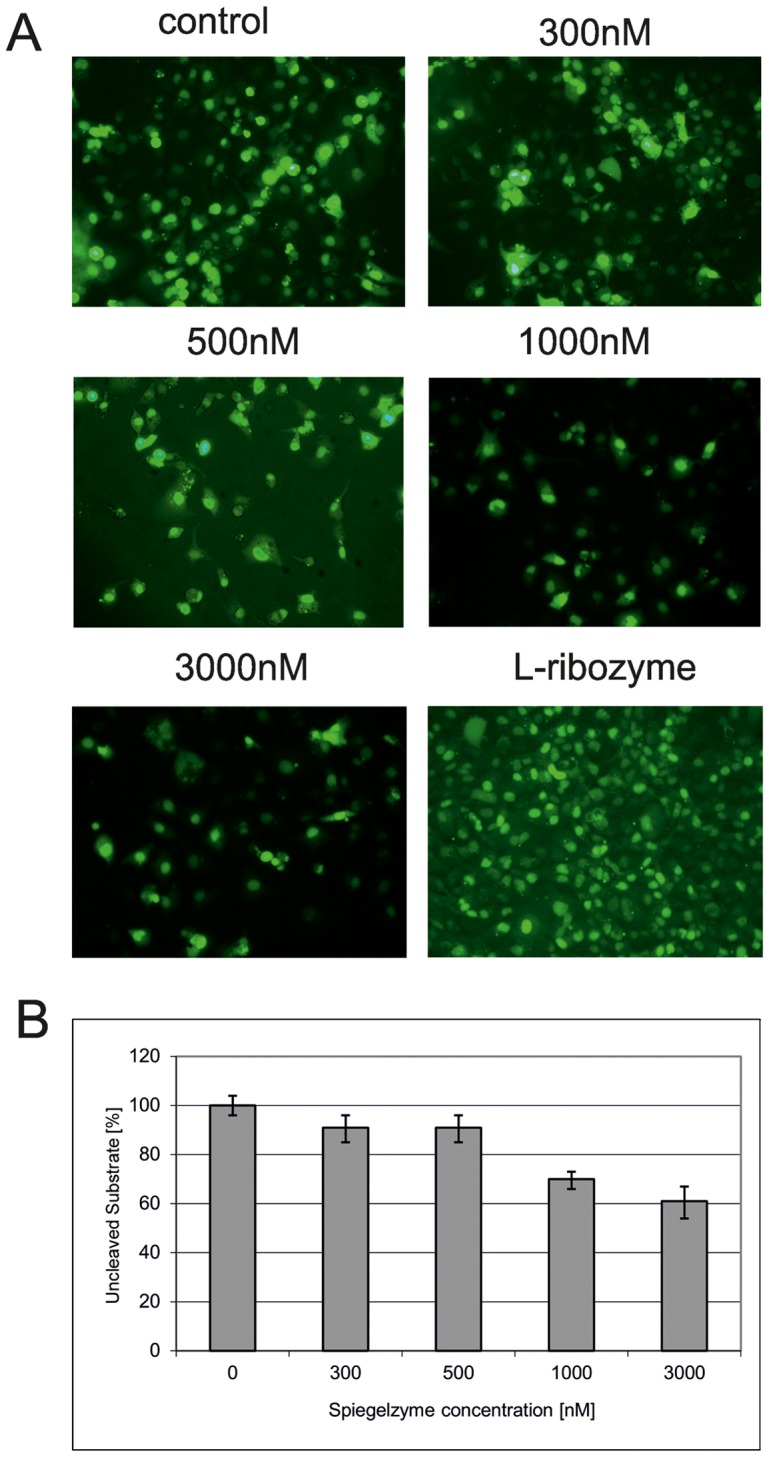
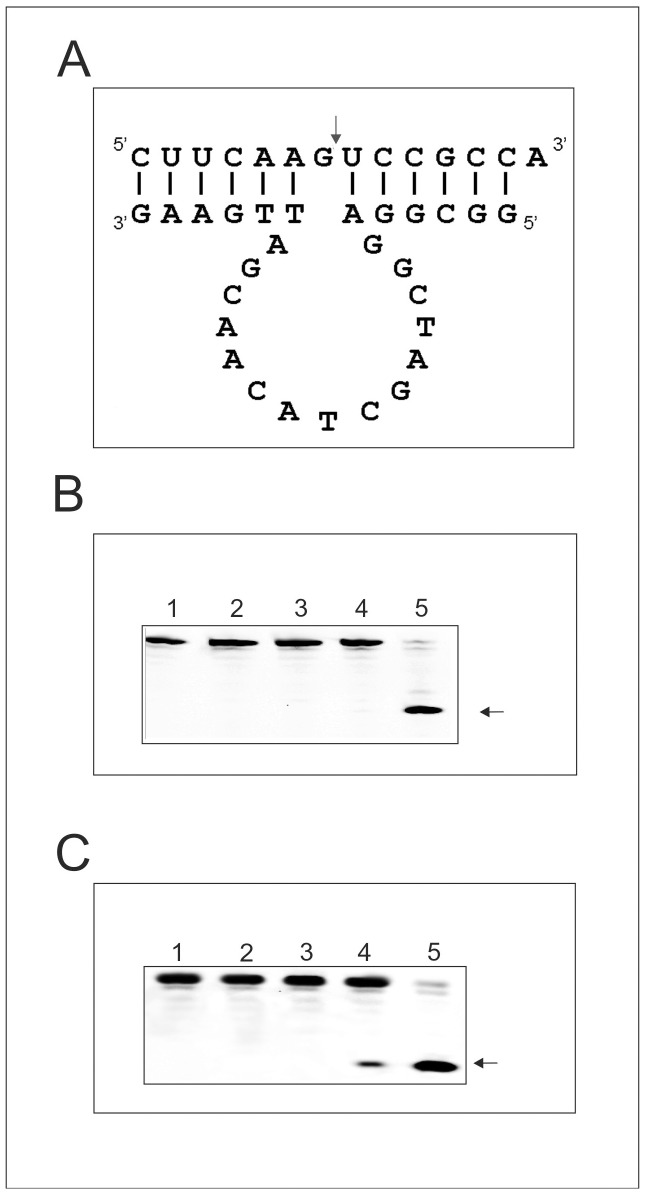
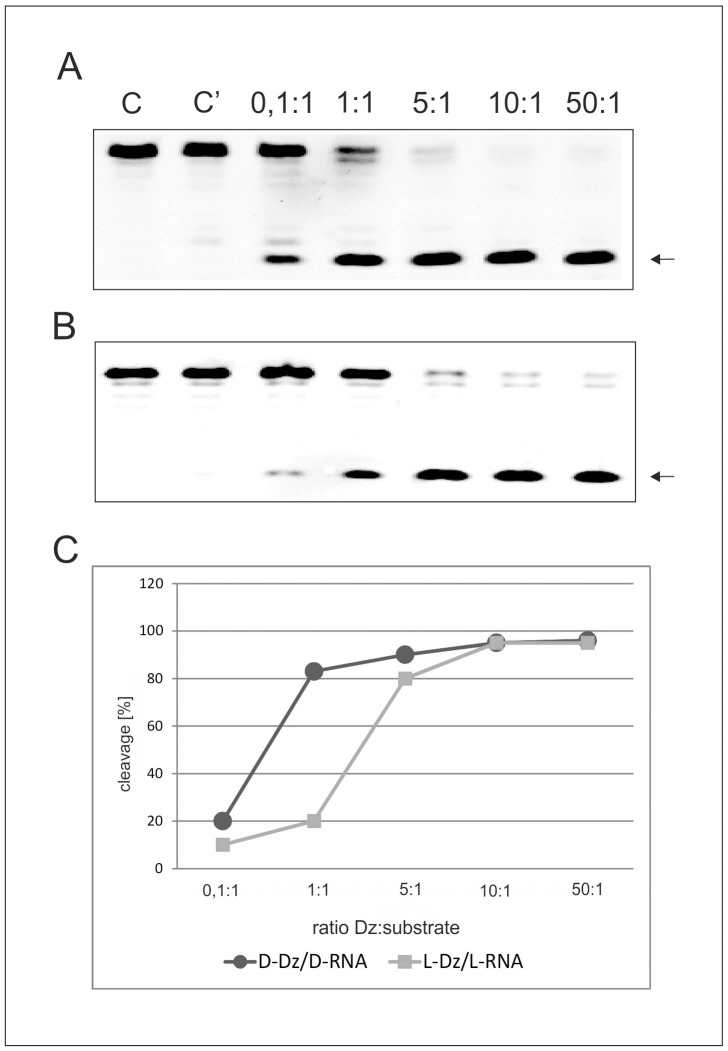
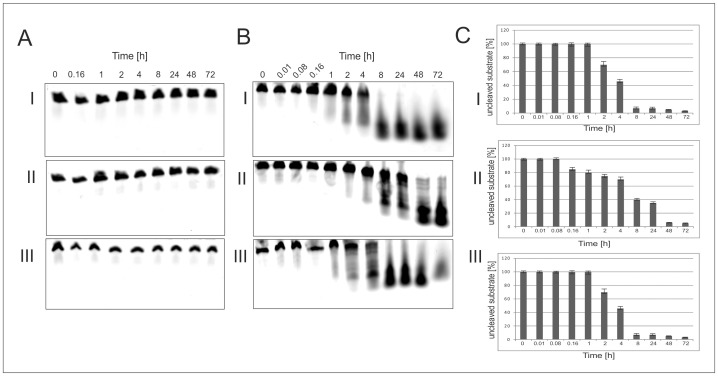
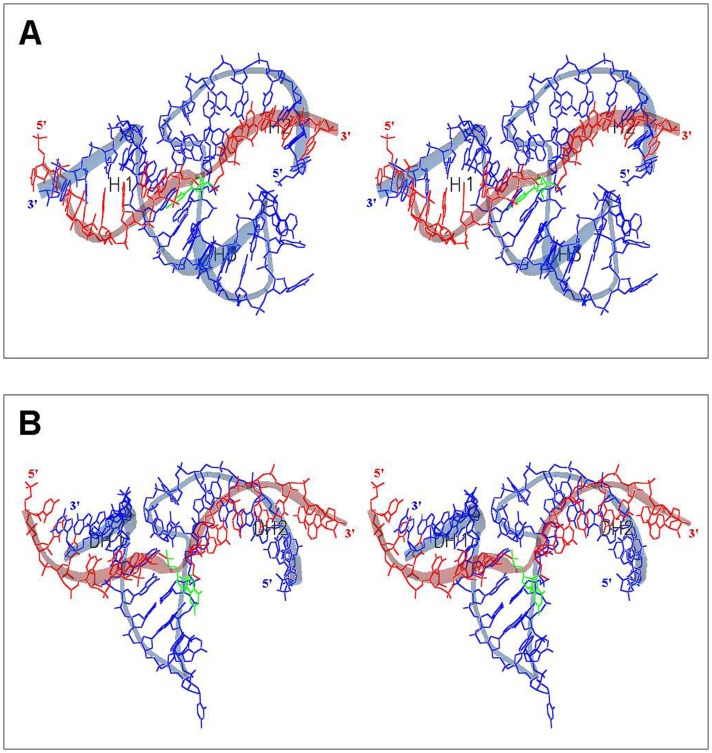
In this manuscript we describe for the first time mirror image catalytic nucleic acids (Spiegelzymes), which hydrolyze sequence specifically L-ribonucleic acid molecules. The mirror image nucleic acid ribozymes designed are based upon the known hammerhead ribozyme and DNAzyme structures that contain L-ribose or L-deoxyribose instead of the naturally occurring D-ribose or D-deoxyribose, respectively. Both Spiegelzymes show similar hydrolytic activities with the same L-RNA target molecules and they also exhibit extra ordinary stabilities when tested with three different human sera. In this respect they are very similar to Spiegelmers (mirror image aptamers), which we had previously developed and for which it has been shown that they are non-toxic and non-immunogenic. Since we are also able to demonstrate that the hammerhead and DNAzyme Spiegelzymes can also hydrolyze mirror image oligonucleotide sequences, like they occur in Spiegelmers, in vivo, it seems reasonable to assume that Spiegelzymes may in principle be used as an antidote against Spiegelmers. Since the Spiegelzymes contain the same building blocks as the Spiegelmers, it can be expected that they will have similar favorable biological characteristics concerning toxicity and immunogenety. In trying to understand the mechanism of action of the Spiegelzymes described in this study, we have initiated for the first time a model building system with L-nucleic acids. The models for L-hammerhead ribozyme and L-DNAzyme interaction with the same L-RNA target will be presented.
Conflict of interest statement
Figures
References
-
- Carothers JM, Szostak JW (2006) In vitro selection of functional oligonucleotides and the origins of biochemical activity. In: The Aptamer Handbook, Functional Oligonuleotides and Their Applications, Ed. S. Klussmann, Wiley-VCH Weinheim.
-
- Klussmann S, Nolte A, Bald R, Erdmann VA, Fürste JP (1996) Mirror-image RNA that binds D-adenosine. Nat Biotechnol 14: 1112–1115. - PubMed
-
- Nolte A, Klussmann S, Bald R, Erdmann VA, Fürste JP (1996) Mirror-design of L-oligonucleotide ligands binding to L-arginine. Nat Biotechnol 14: 1116–1119. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
