

Reduced interleukin-4 receptor α expression on CD8+ T cells correlates with higher quality anti-viral immunity
- PMID: 23383283
- PMCID: PMC3561338
- DOI: 10.1371/journal.pone.0055788
Reduced interleukin-4 receptor α expression on CD8+ T cells correlates with higher quality anti-viral immunity
Abstract
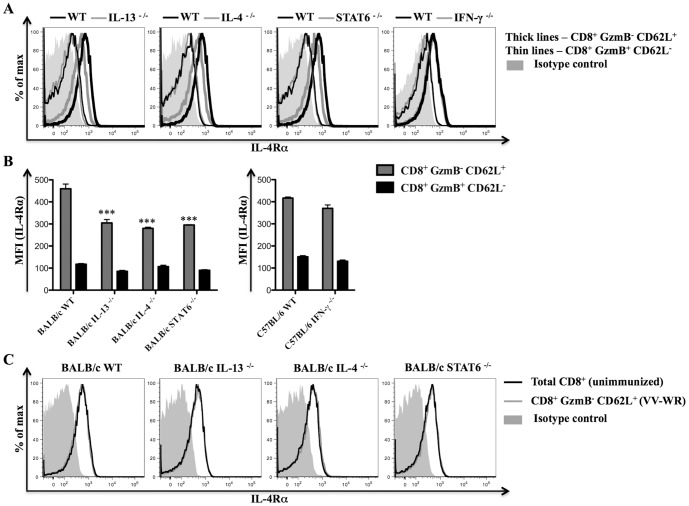
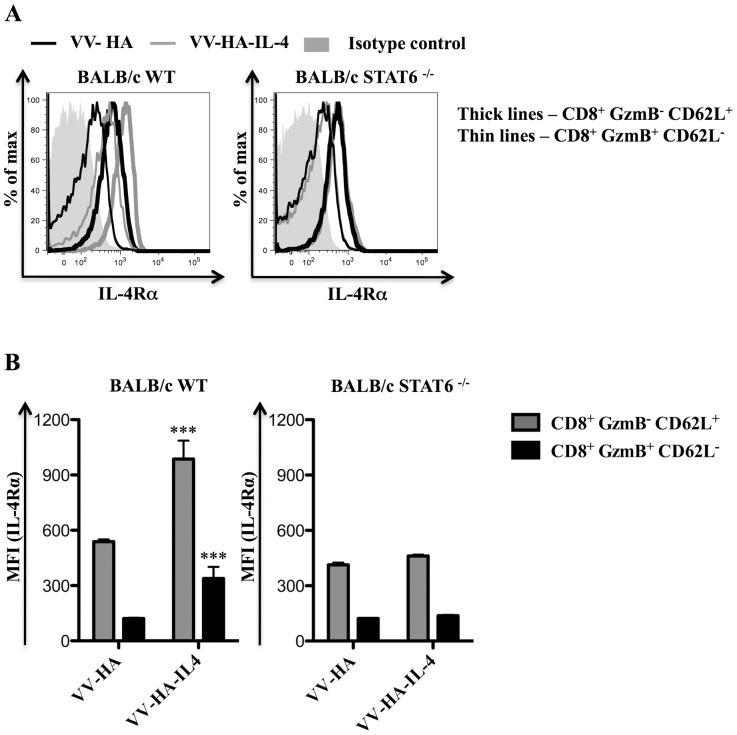
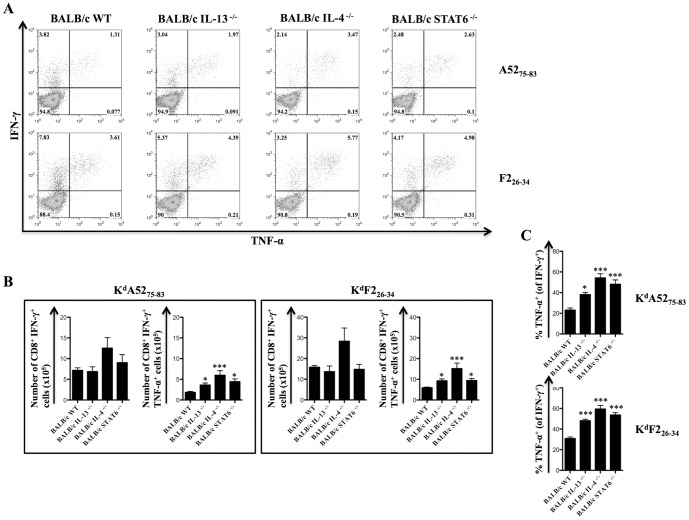
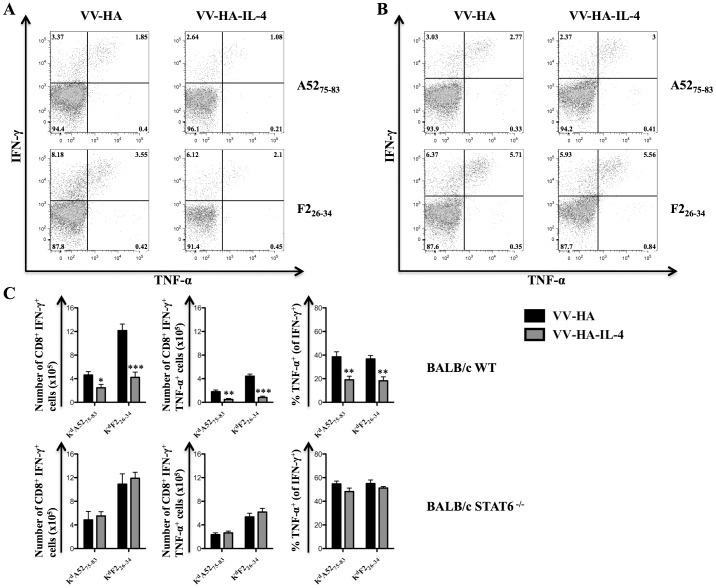
With the hope of understanding how interleukin (IL)-4 and IL-13 modulated quality of anti-viral CD8(+) T cells, we evaluated the expression of receptors for these cytokines following a range of viral infections (e.g. pox viruses and influenza virus). Results clearly indicated that unlike other IL-4/IL-13 receptor subunits, IL-4 receptor α (IL-4Rα) was significantly down-regulated on anti-viral CD8(+) T cells in a cognate antigen dependent manner. The infection of gene knockout mice and wild-type (WT) mice with vaccinia virus (VV) or VV expressing IL-4 confirmed that IL-4, IL-13 and signal transducer and activator of transcription 6 (STAT6) were required to increase IL-4Rα expression on CD8(+) T cells, but not interferon (IFN)-γ. STAT6 dependent elevation of IL-4Rα expression on CD8(+) T cells was a feature of poor quality anti-viral CD8(+) T cell immunity as measured by the production of IFN-γ and tumor necrosis factor α (TNF-α) in response to VV antigen stimulation in vitro. We propose that down-regulation of IL-4Rα, but not the other IL-4/IL-13 receptor subunits, is a mechanism by which CD8(+) T cells reduce responsiveness to IL-4 and IL-13. This can improve the quality of anti-viral CD8(+) T cell immunity. Our findings have important implications in understanding anti-viral CD8(+) T cell immunity and designing effective vaccines against chronic viral infections.
Conflict of interest statement
Figures
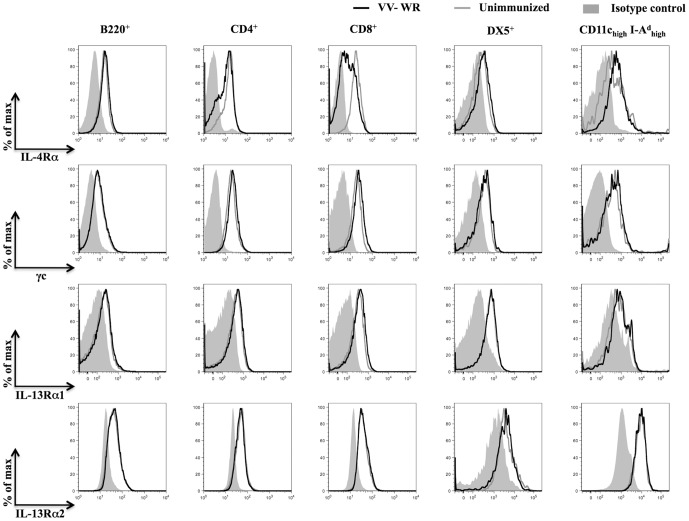
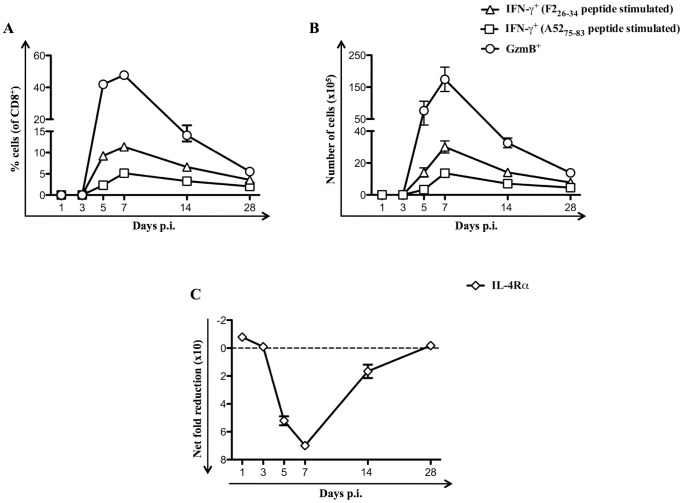
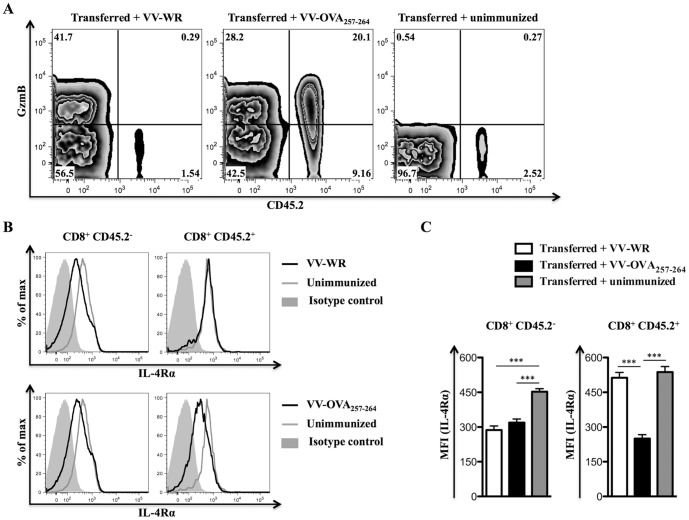
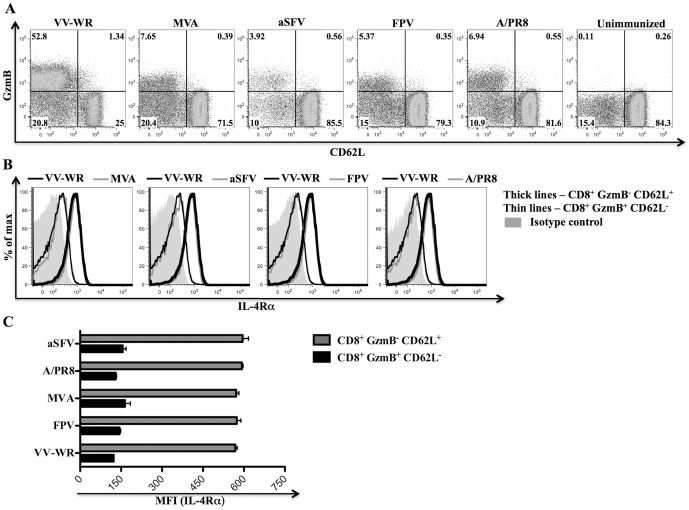
References
-
- Harty JT, Tvinnereim AR, White DW (2000) CD8+ T cell effector mechanisms in resistance to infection. Annu Rev Immunol 18: 275–308. - PubMed
-
- Zinkernagel RM, Doherty PC (1974) Restriction of in vitro T cell-mediated cytotoxicity in lymphocytic choriomeningitis within a syngeneic or semiallogeneic system. Nature 248: 701–702. - PubMed
-
- Seder RA, Darrah PA, Roederer M (2008) T-cell quality in memory and protection: implications for vaccine design. Nat Rev Immunol 8: 247–258. - PubMed
-
- Derby M, Alexander-Miller M, Tse R, Berzofsky J (2001) High-avidity CTL exploit two complementary mechanisms to provide better protection against viral infection than low-avidity CTL. J Immunol 166: 1690–1697. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
