

Doublecortin-like kinase enhances dendritic remodelling and negatively regulates synapse maturation
- PMID: 23385585
- PMCID: PMC4017031
- DOI: 10.1038/ncomms2443
Doublecortin-like kinase enhances dendritic remodelling and negatively regulates synapse maturation
Abstract
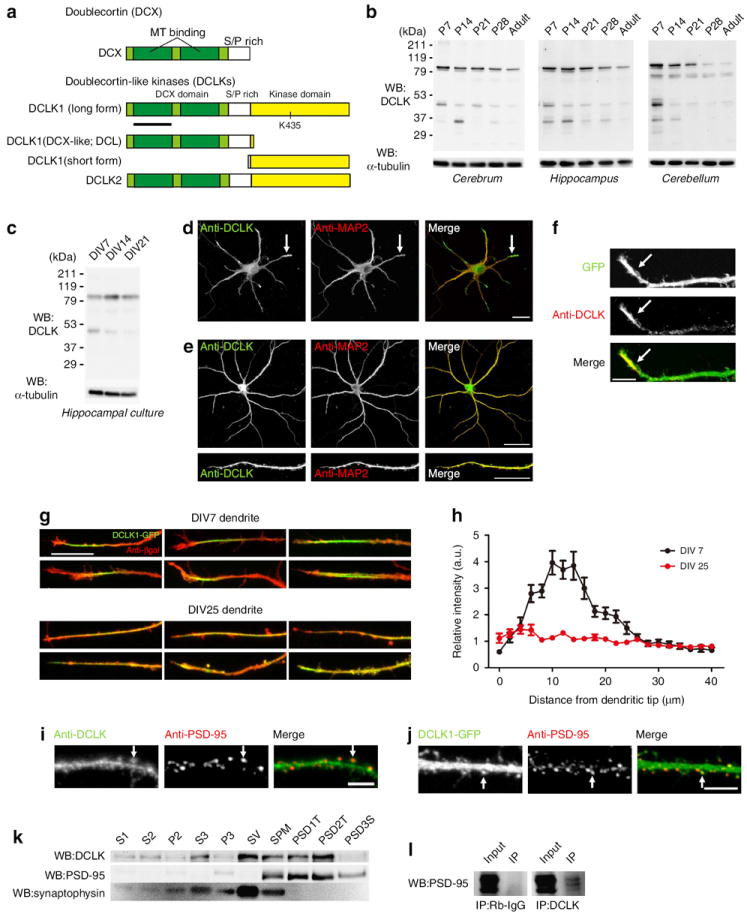
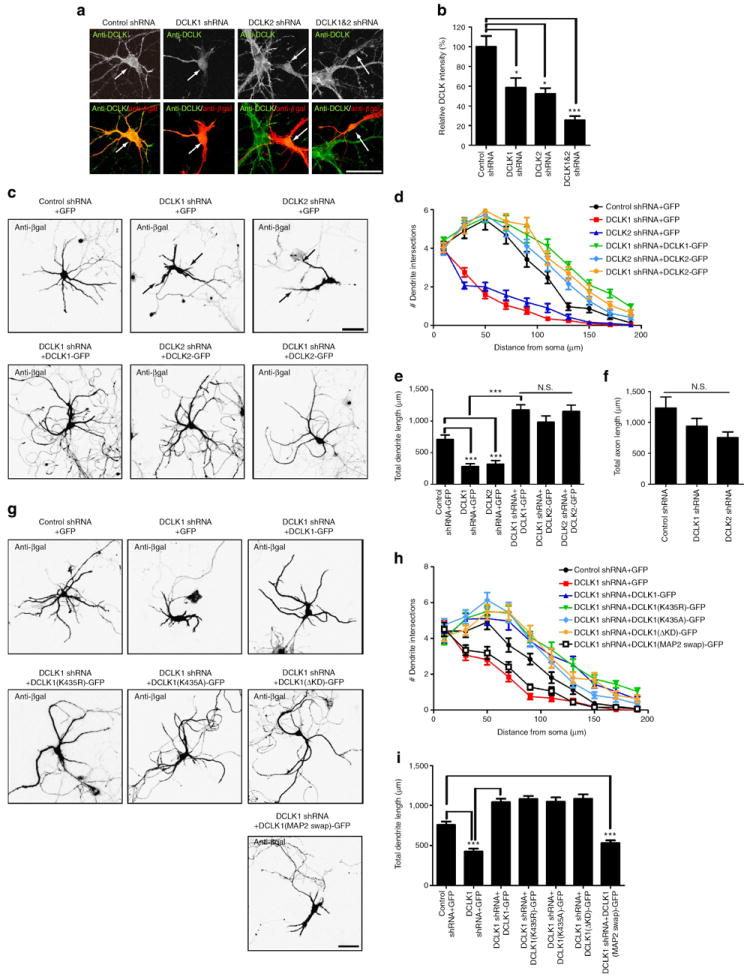
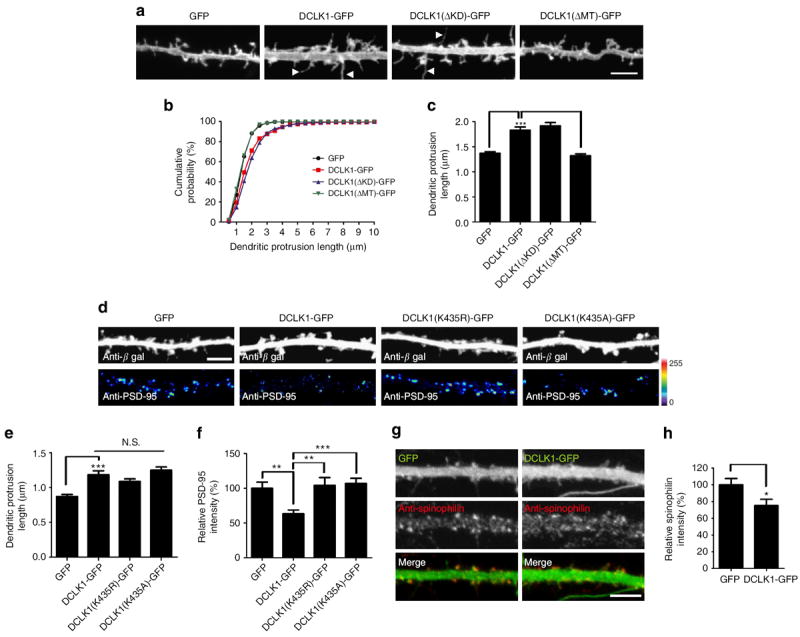
Dendritic morphogenesis and formation of synapses at appropriate dendritic locations are essential for the establishment of proper neuronal connectivity. Recent imaging studies provide evidence for stabilization of dynamic distal branches of dendrites by the addition of new synapses. However, molecules involved in both dendritic growth and suppression of synapse maturation remain to be identified. Here we report two distinct functions of doublecortin-like kinases, chimeric proteins containing both a microtubule-binding domain and a kinase domain in postmitotic neurons. First, doublecortin-like kinases localize to the distal dendrites and promote their growth by enhancing microtubule bundling. Second, doublecortin-like kinases suppress maturation of synapses through multiple pathways, including reduction of PSD-95 by the kinase domain and suppression of spine structural maturation by the microtubule-binding domain. Thus, doublecortin-like kinases are critical regulators of dendritic development by means of their specific targeting to the distal dendrites, and their local control of dendritic growth and synapse maturation.
Conflict of interest statement
Figures




References
-
- Allen KM, Walsh CA. Genes that regulate neuronal migration in the cerebral cortex. Epilepsy Res. 1999;36:143–154. - PubMed
-
- Gleeson JG, et al. Doublecortin, a brain-specific gene mutated in human X-linked lissencephaly and double cortex syndrome, encodes a putative signaling protein. Cell. 1998;92:63–72. - PubMed
-
- Taylor KR, Holzer AK, Bazan JF, Walsh CA, Gleeson JG. Patient mutations in doublecortin define a repeated tubulin-binding domain. J Biol Chem. 2000;275:34442–34450. - PubMed
-
- Francis F, et al. Doublecortin is a developmentally regulated, microtubule-associated protein expressed in migrating and differentiating neurons. Neuron. 1999;23:247–256. - PubMed
-
- Gleeson JG, Lin PT, Flanagan LA, Walsh CA. Doublecortin is a microtubule-associated protein and is expressed widely by migrating neurons. Neuron. 1999;23:257–271. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
