

Kissing and nanotunneling mediate intermitochondrial communication in the heart
- PMID: 23386722
- PMCID: PMC3581932
- DOI: 10.1073/pnas.1300741110
Kissing and nanotunneling mediate intermitochondrial communication in the heart
Abstract
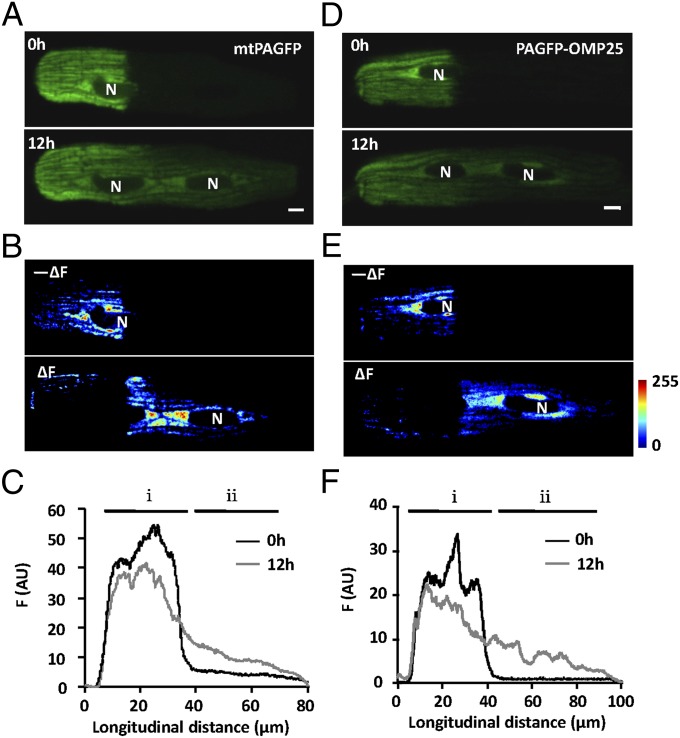
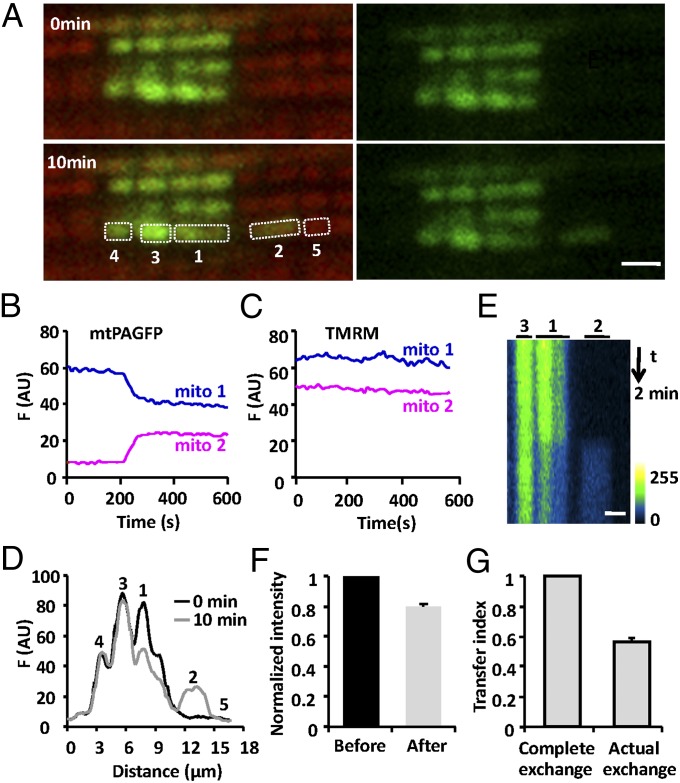
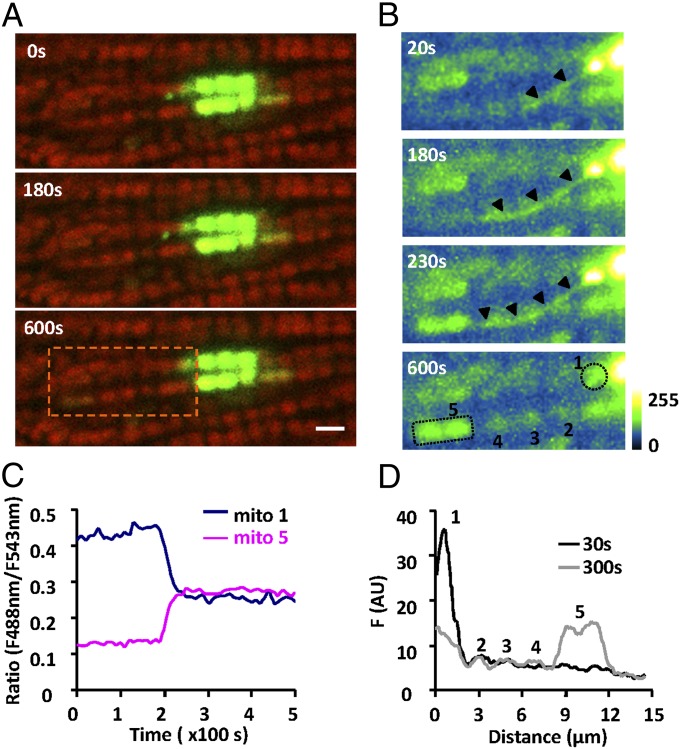
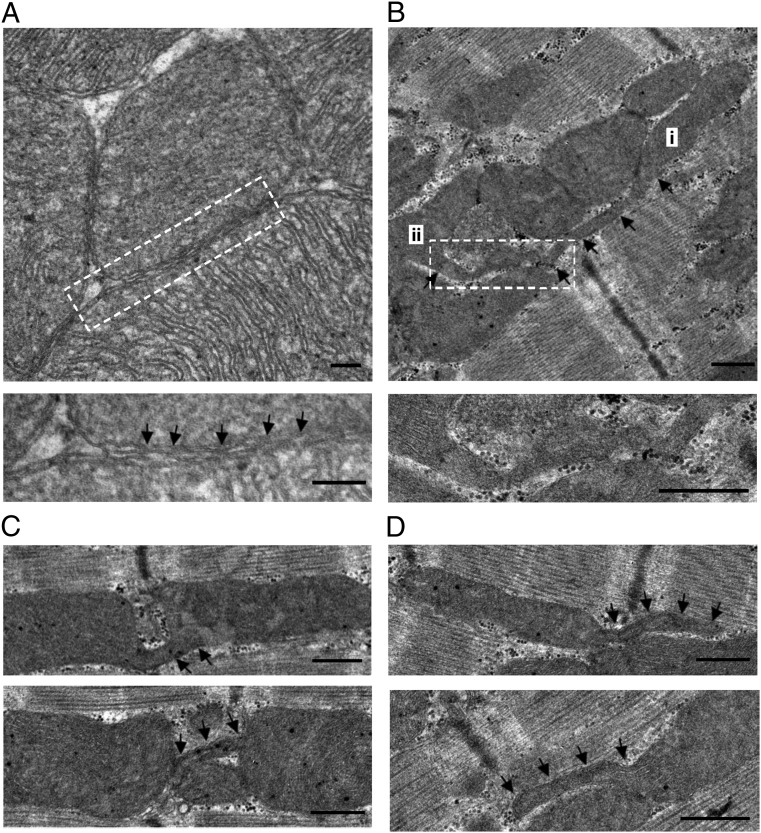
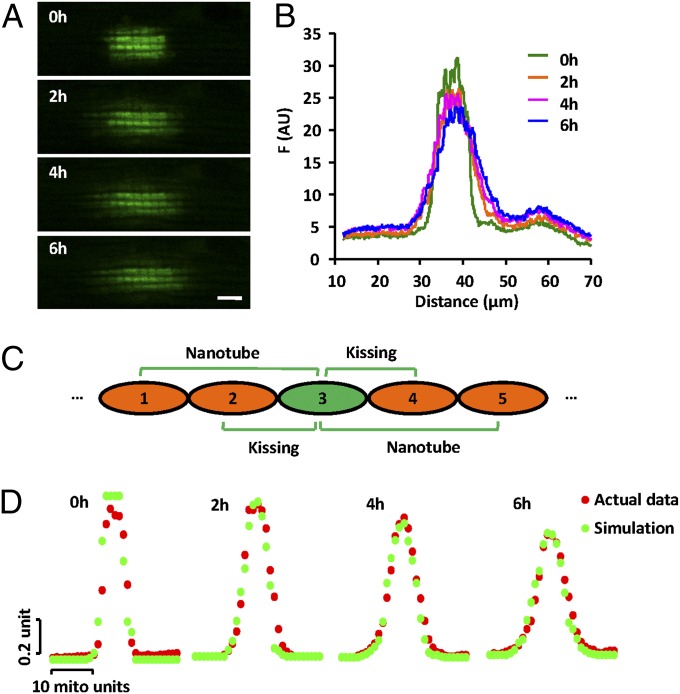
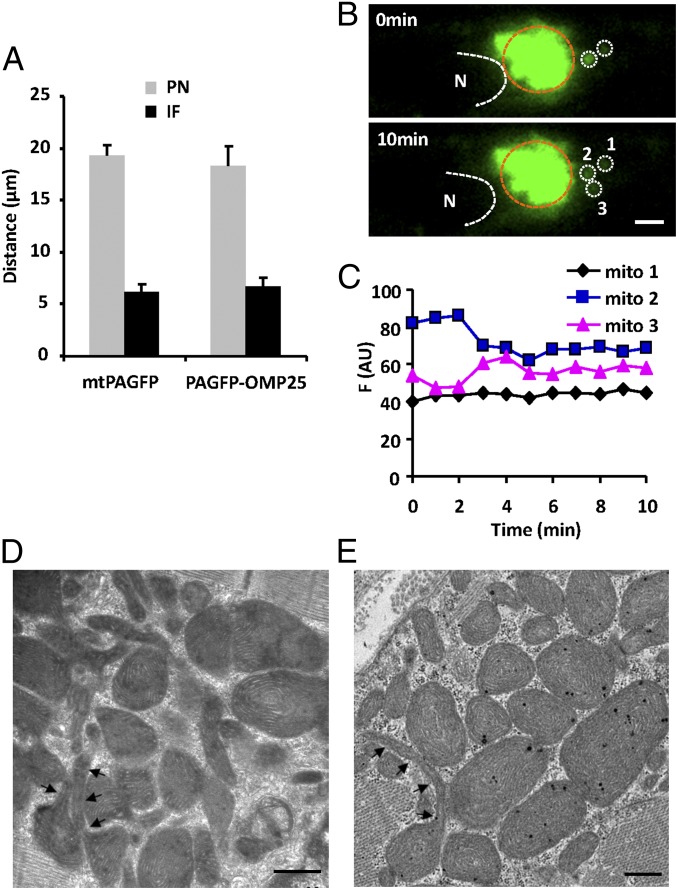
Mitochondria in many types of cells are dynamically interconnected through constant fusion and fission, allowing for exchange of mitochondrial contents and repair of damaged mitochondria. However, constrained by the myofibril lattice, the ∼6,000 mitochondria in the adult mammalian cardiomyocyte display little motility, and it is unclear how, if at all, they communicate with each other. By means of target-expressing photoactivatable green fluorescent protein (PAGFP) in the mitochondrial matrix or on the outer mitochondrial membrane, we demonstrated that the local PAGFP signal propagated over the entire population of mitochondria in cardiomyocytes on a time scale of ∼10 h. Two elemental steps of intermitochondrial communications were manifested as either a sudden PAGFP transfer between a pair of adjacent mitochondria (i.e., "kissing") or a dynamic nanotubular tunnel (i.e., "nanotunneling") between nonadjacent mitochondria. The average content transfer index (fractional exchange) was around 0.5; the rate of kissing was 1‰ s(-1) per mitochondrial pair, and that of nanotunneling was about 14 times smaller. Electron microscopy revealed extensive intimate contacts between adjacent mitochondria and elongated nanotubular protrusions, providing a structural basis for the kissing and nanotunneling, respectively. We propose that, through kissing and nanotunneling, the otherwise static mitochondria in a cardiomyocyte form one dynamically continuous network to share content and transfer signals.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
