

Borrelia host adaptation Regulator (BadR) regulates rpoS to modulate host adaptation and virulence factors in Borrelia burgdorferi
- PMID: 23387366
- PMCID: PMC4828661
- DOI: 10.1111/mmi.12171
Borrelia host adaptation Regulator (BadR) regulates rpoS to modulate host adaptation and virulence factors in Borrelia burgdorferi
Abstract
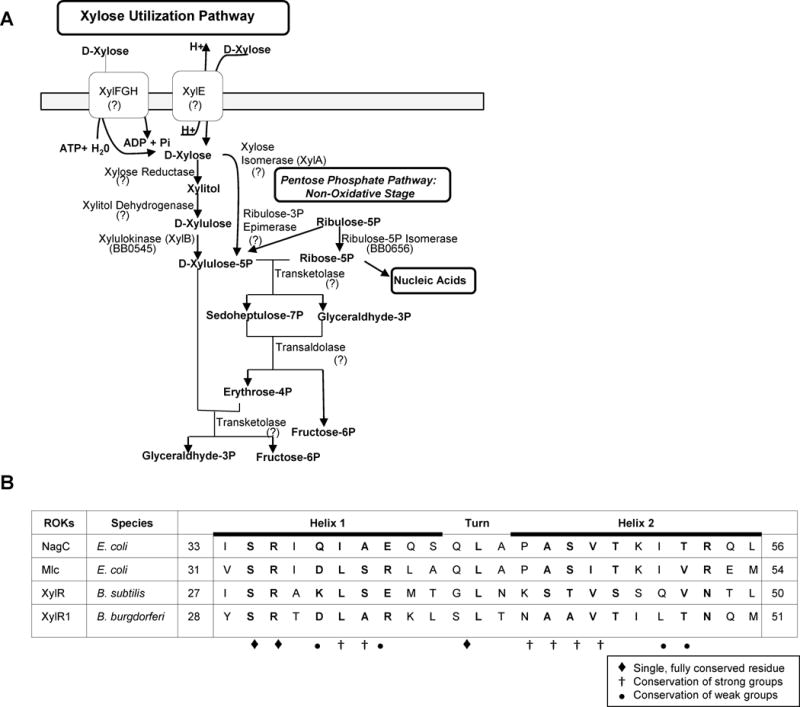
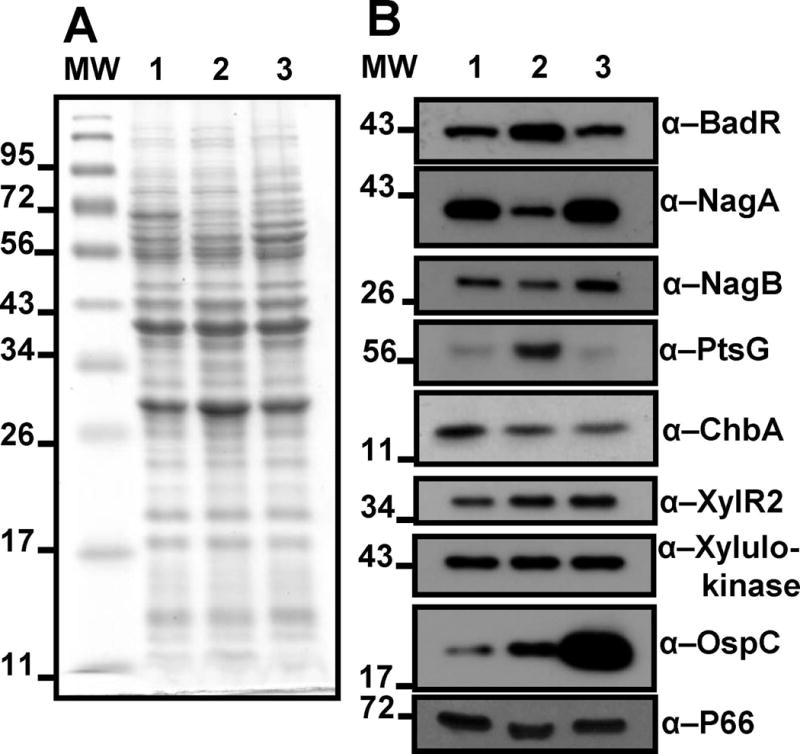
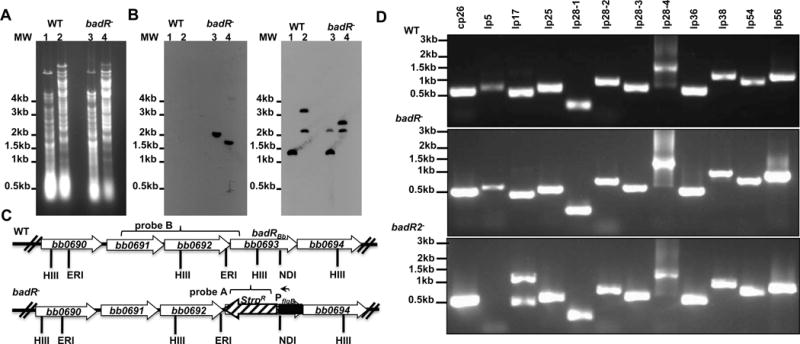
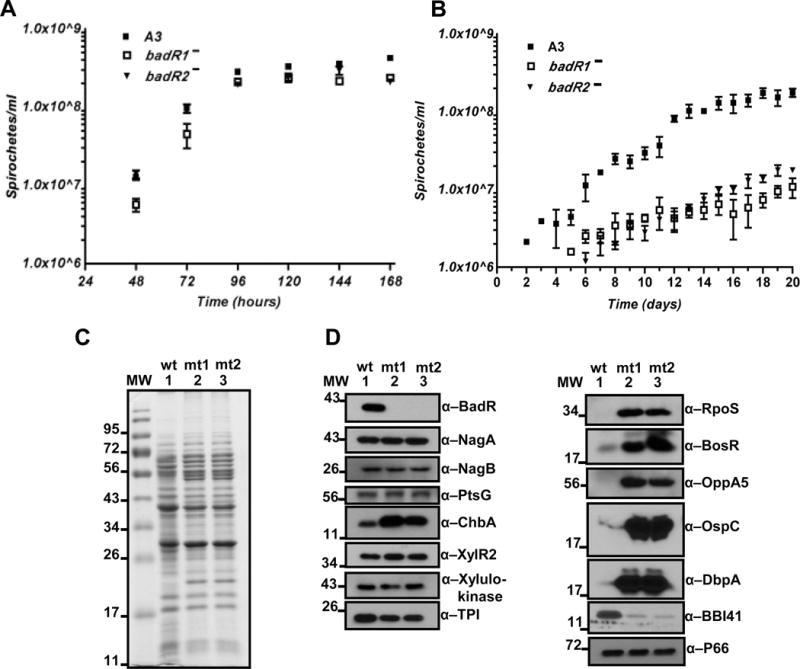
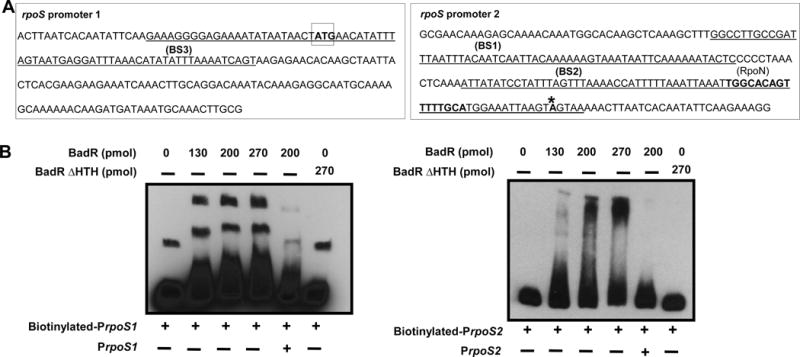
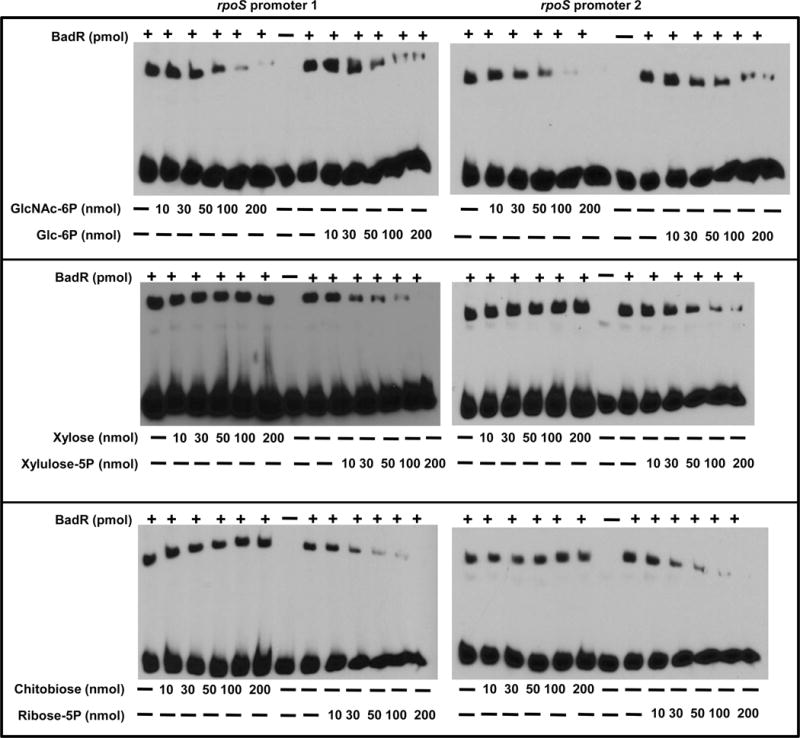
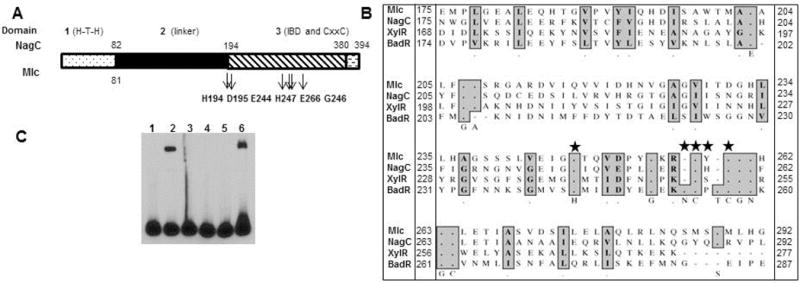
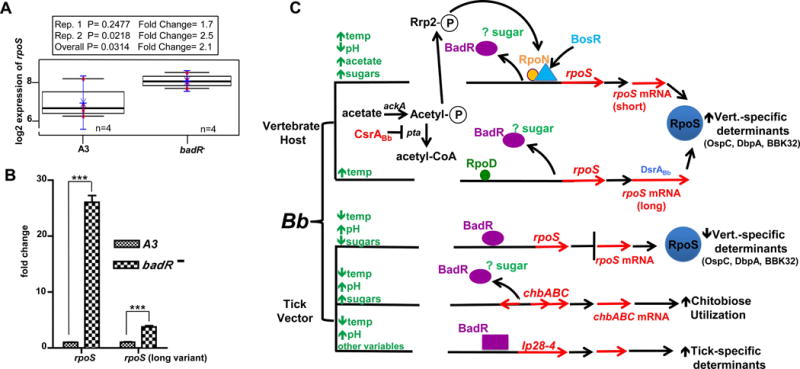
The RpoS transcription factor of Borrelia burgdorferi is a 'gatekeeper' because it activates genes required for spirochaetes to transition from tick to vertebrate hosts. However, it remains unknown how RpoS becomes repressed to allow the spirochaetes to transition back from the vertebrate host to the tick vector. Here we show that a putative carbohydrate-responsive regulatory protein, designated BadR (Borrelia host adaptation Regulator), is a transcriptional repressor of rpoS. BadR levels are elevated in B. burgdorferi cultures grown under in vitro conditions mimicking unfed-ticks and badR-deficient strains are defective for growth under these same conditions. Microarray and immunoblot analyses of badR-deficient strains showed upregulation of rpoS and other factors important for virulence in vertebrate hosts, as well as downregulation of putative tick-specific determinants (e.g. linear plasmid 28-4 genes). DNA-binding assays revealed BadR binds to upstream regions of rpoS. Site-directed mutations in BadR and the presence of phosphorylated sugars affected BadR's binding to the rpoS promoters. badR-deficient B. burgdorferi were unable to colonize mice. Several putative tick-specific targets have been identified. Our study identified a novel regulator, BadR, and provides a link between nutritional environmental cues utilized by spirochaetes to adaptation to disparate conditions found in the tick and vertebrate hosts.
© 2013 Blackwell Publishing Ltd.
Figures
References
-
- Berg T, Schild S, Reidl J. Regulation of the chitobiose-phosphotransferase system in Vibrio cholerae. Arch Microbiol. 2007;187:433–439. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
