

The role of BRCA1 in homologous recombination repair in response to replication stress: significance in tumorigenesis and cancer therapy
- PMID: 23388117
- PMCID: PMC3599463
- DOI: 10.1186/2045-3701-3-11
The role of BRCA1 in homologous recombination repair in response to replication stress: significance in tumorigenesis and cancer therapy
Abstract
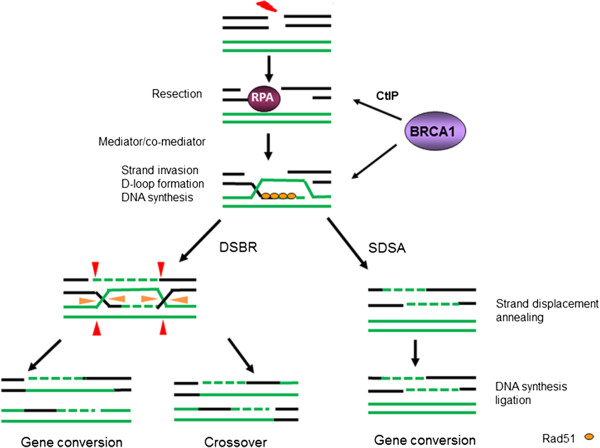
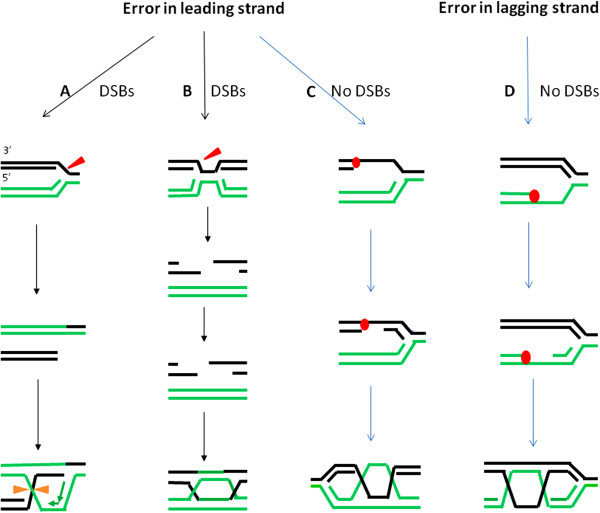
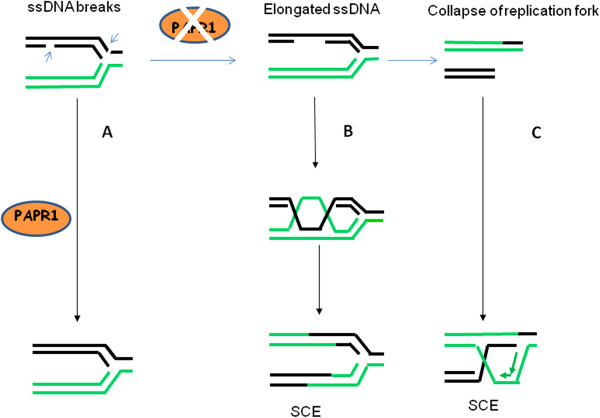
Germ line mutations in breast cancer gene 1 (BRCA1) predispose women to breast and ovarian cancers. Although BRCA1 is involved in many important biological processes, the function of BRCA1 in homologous recombination (HR) mediated repair is considered one of the major mechanisms contributing to its tumor suppression activity, and the cause of hypersensitivity to poly(ADP-ribose) polymerase (PARP) inhibitors when BRCA1 is defective. Mounting evidence suggests that the mechanism of repairing DNA double strand breaks (DSBs) by HR is different than the mechanism operating when DNA replication is blocked. Although BRCA1 has been recognized as a central component in HR, the precise role of BRCA1 in HR, particularly under replication stress, has remained largely unknown. Given the fact that DNA lesions caused by replication blockages are the primary substrates for HR in mitotic cells, functional analysis of BRCA1 in HR repair in the context of replication stress should benefit our understanding of the molecular mechanisms underlying tumorigenesis associated with BRCA1 deficiencies, as well as the development of therapeutic approaches for cancer patients carrying BRCA1 mutations or reduced BRCA1 expression. This review focuses on the current advances in this setting and also discusses the significance in tumorigenesis and cancer therapy.
Figures
References
-
- Assenmacher N, Hopfner KP. MRE11/RAD50/NBS1: complex activities. Chromosoma. 2004;113(4):157–166. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
