

Splenic priming of virus-specific CD8 T cells following influenza virus infection
- PMID: 23388712
- PMCID: PMC3624369
- DOI: 10.1128/JVI.03413-12
Splenic priming of virus-specific CD8 T cells following influenza virus infection
Abstract
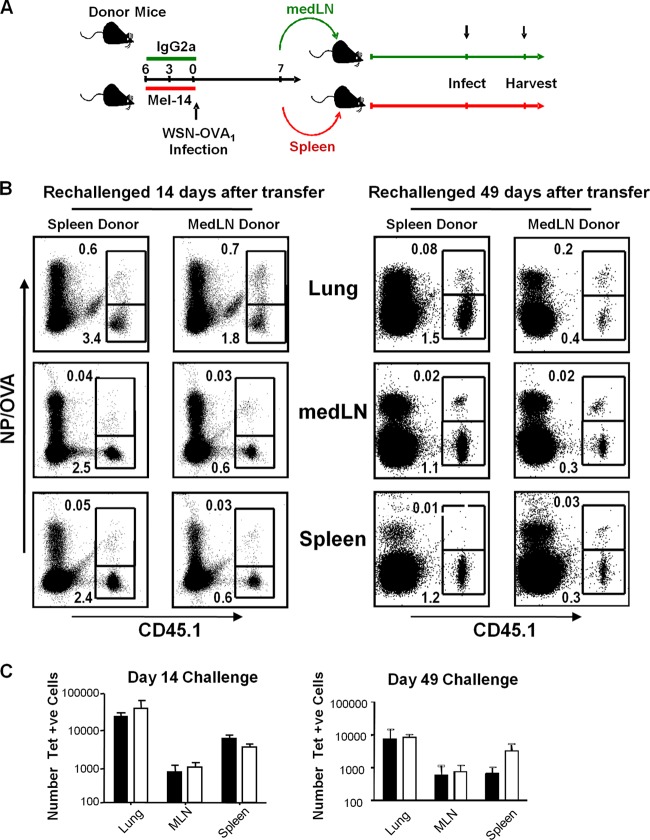
In healthy individuals, influenza virus (IAV) infection generally remains localized to the epithelial cells of the respiratory tract. Previously, IAV-specific effector CD8 T cells found systemically during the course of IAV infection were thought to have been primed in lung-draining lymph nodes with subsequent migration to other tissues. However, little is known about whether other lymphoid sites participate in the generation of virus-specific CD8 T cells during localized IAV infection. Here, we present evidence of early CD8 T cell priming in the spleen following respiratory IAV infection independent of lung-draining lymph node priming of T cells. Although we found early indications of CD8 T cell activation in the lymph nodes draining the respiratory tract, we also saw evidence of virus-specific CD8 T cell activation in the spleen. Furthermore, CD8 T cells primed in the spleen differentiated into memory cells of equivalent longevity and with similar recall capacity as CD8 T cells primed in the draining lymph nodes. These data showed that the spleen contributes to the virus-specific effector and memory CD8 T cell populations that are generated in response to respiratory infection.
Figures






Similar articles
-
Modeling of influenza-specific CD8+ T cells during the primary response indicates that the spleen is a major source of effectors.J Immunol. 2011 Nov 1;187(9):4474-82. doi: 10.4049/jimmunol.1101443. Epub 2011 Sep 23. J Immunol. 2011. PMID: 21948988 Free PMC article.
-
During viral infection of the respiratory tract, CD27, 4-1BB, and OX40 collectively determine formation of CD8+ memory T cells and their capacity for secondary expansion.J Immunol. 2005 Aug 1;175(3):1665-76. doi: 10.4049/jimmunol.175.3.1665. J Immunol. 2005. PMID: 16034107
-
Immunity to Respiratory Infection Is Reinforced Through Early Proliferation of Lymphoid TRM Cells and Prompt Arrival of Effector CD8 T Cells in the Lungs.Front Immunol. 2019 Jun 14;10:1370. doi: 10.3389/fimmu.2019.01370. eCollection 2019. Front Immunol. 2019. PMID: 31258537 Free PMC article.
-
Characterization of CD8+ T cell repertoire diversity and persistence in the influenza A virus model of localized, transient infection.Semin Immunol. 2004 Jun;16(3):179-84. doi: 10.1016/j.smim.2004.02.005. Semin Immunol. 2004. PMID: 15130502 Review.
-
Primary and long-term B-cell responses in the upper airway and lung after influenza A virus infection.Immunol Res. 2014 Aug;59(1-3):73-80. doi: 10.1007/s12026-014-8541-0. Immunol Res. 2014. PMID: 24838149 Review.
Cited by
-
Identifying Active Compounds and Targets of Fritillariae thunbergii against Influenza-Associated Inflammation by Network Pharmacology Analysis and Molecular Docking.Molecules. 2020 Aug 25;25(17):3853. doi: 10.3390/molecules25173853. Molecules. 2020. PMID: 32854331 Free PMC article.
-
Immunity to Influenza Infection in Humans.Cold Spring Harb Perspect Med. 2021 Mar 1;11(3):a038729. doi: 10.1101/cshperspect.a038729. Cold Spring Harb Perspect Med. 2021. PMID: 31871226 Free PMC article. Review.
-
T cell kinetics reveal expansion of distinct lung T cell subsets in acute versus in resolved influenza virus infection.Front Immunol. 2022 Oct 6;13:949299. doi: 10.3389/fimmu.2022.949299. eCollection 2022. Front Immunol. 2022. PMID: 36275685 Free PMC article.
-
An imbalance between innate and adaptive immune cells at the maternal-fetal interface occurs prior to endotoxin-induced preterm birth.Cell Mol Immunol. 2016 Jul;13(4):462-73. doi: 10.1038/cmi.2015.22. Epub 2015 Apr 6. Cell Mol Immunol. 2016. PMID: 25849119 Free PMC article.
-
TCR Transgenic Mice: A Valuable Tool for Studying Viral Immunopathogenesis Mechanisms.Int J Mol Sci. 2020 Dec 18;21(24):9690. doi: 10.3390/ijms21249690. Int J Mol Sci. 2020. PMID: 33353154 Free PMC article. Review.
References
-
- Inaba K, Turley S, Yamaide F, Iyoda T, Mahnke K, Inaba M, Pack M, Subklewe M, Sauter B, Sheff D, Albert M, Bhardwaj N, Mellman I, Steinman RM. 1998. Efficient presentation of phagocytosed cellular fragments on the major histocompatibility complex class II products of dendritic cells. J. Exp. Med. 188:2163–2173 - PMC - PubMed
-
- Albert ML, Sauter B, Bhardwaj N. 1998. Dendritic cells acquire antigen from apoptotic cells and induce class I-restricted CTLs. Nature 392:86–89 - PubMed
-
- Bender A, Albert M, Reddy A, Feldman M, Sauter B, Kaplan G, Hellman W, Bhardwaj N. 1998. The distinctive features of influenza virus infection of dendritic cells. Immunobiology 198:552–567 - PubMed
-
- Legge KL, Braciale TJ. 2003. Accelerated migration of respiratory dendritic cells to the regional lymph nodes is limited to the early phase of pulmonary infection. Immunity 18:265–277 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
