

Tetraspanin CD9 links junctional adhesion molecule-A to αvβ3 integrin to mediate basic fibroblast growth factor-specific angiogenic signaling
- PMID: 23389628
- PMCID: PMC3608503
- DOI: 10.1091/mbc.E12-06-0481
Tetraspanin CD9 links junctional adhesion molecule-A to αvβ3 integrin to mediate basic fibroblast growth factor-specific angiogenic signaling
Abstract
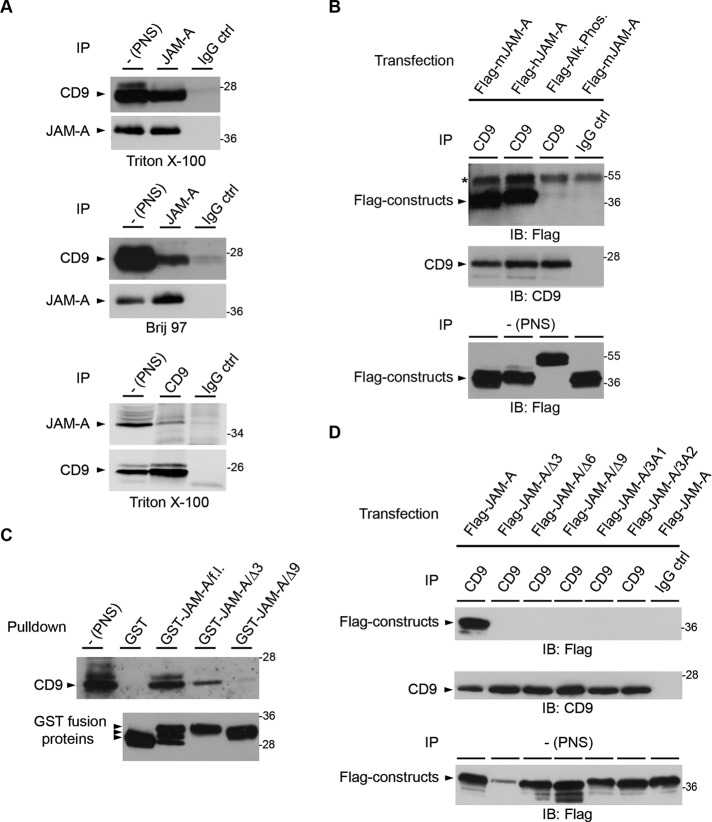
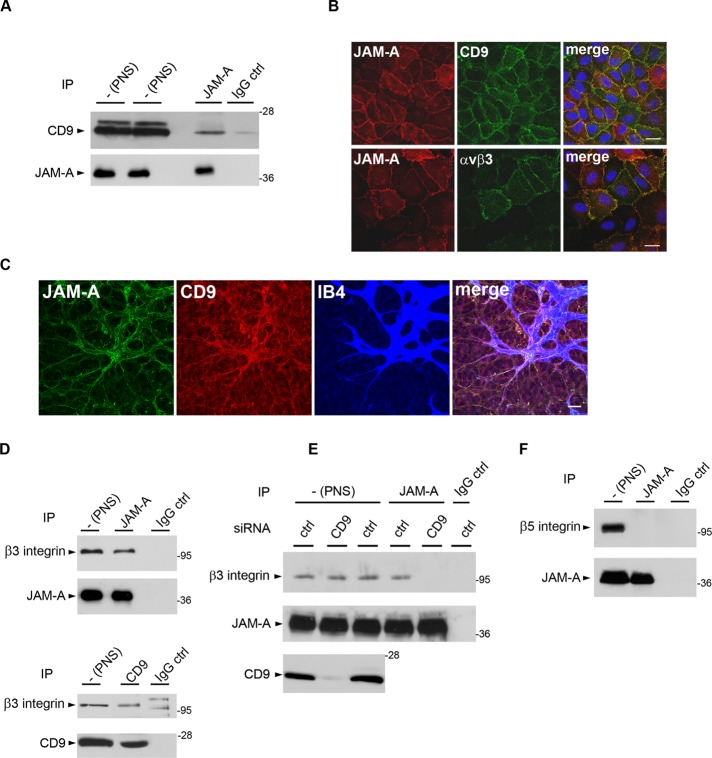
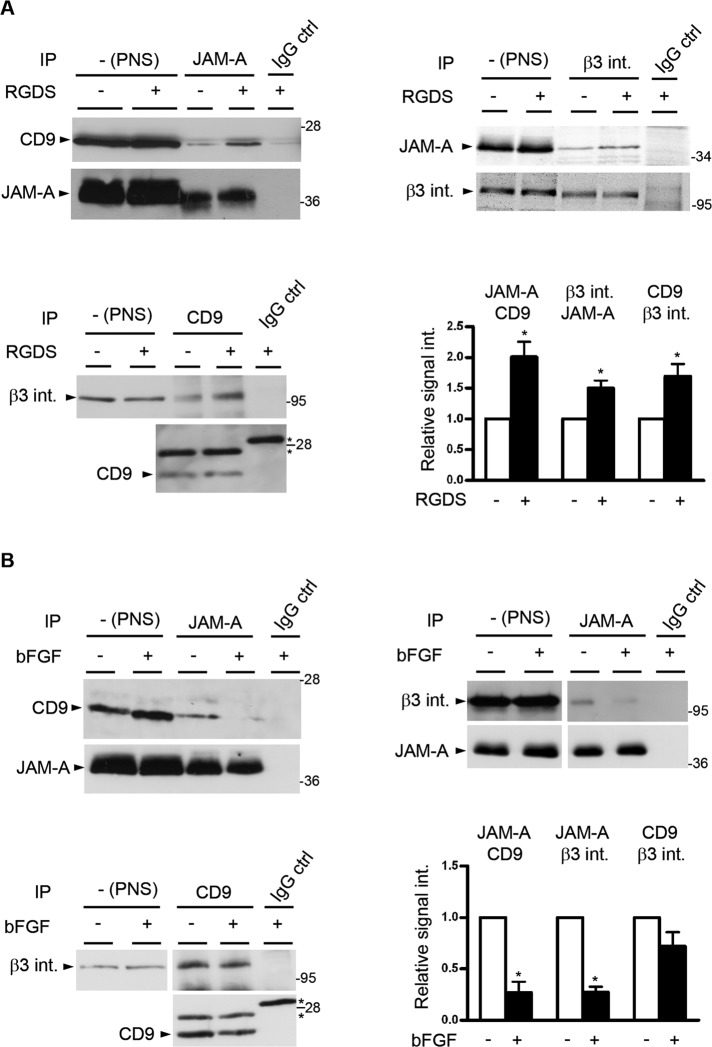
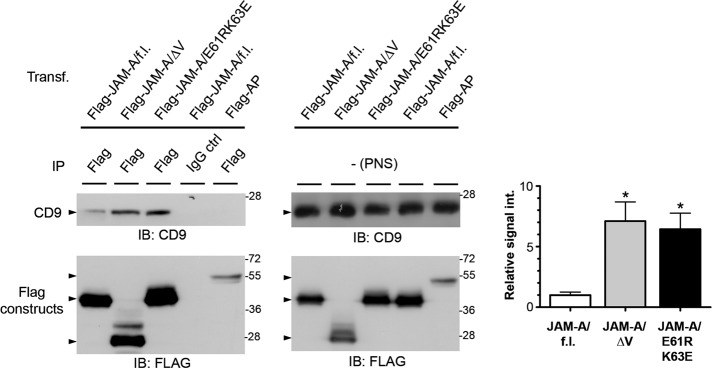
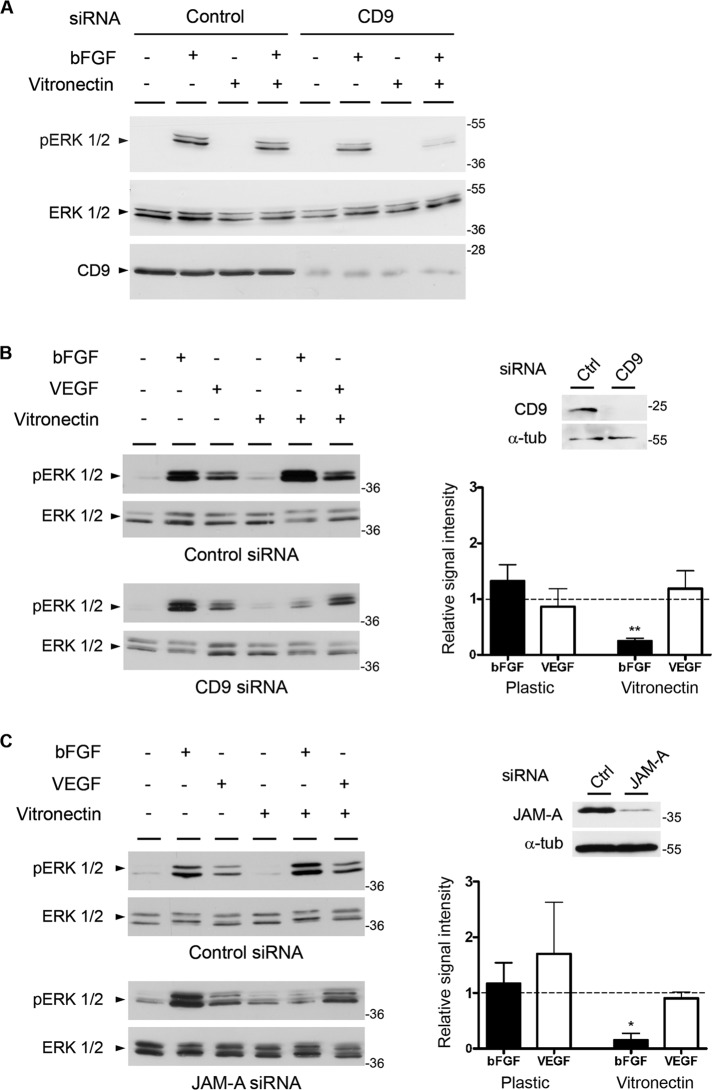
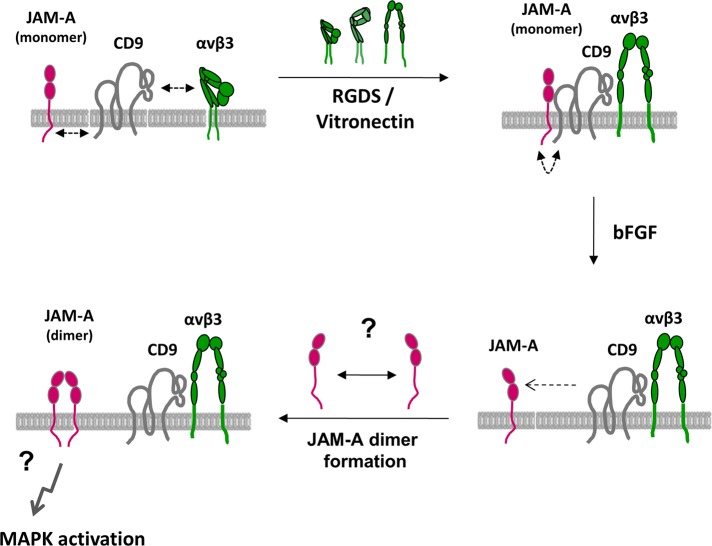
Junctional adhesion molecule-A (JAM-A) is a member of the immunoglobulin family with diverse functions in epithelial cells, including cell migration, cell contact maturation, and tight junction formation. In endothelial cells, JAM-A has been implicated in basic fibroblast growth factor (bFGF)-regulated angiogenesis through incompletely understood mechanisms. In this paper, we identify tetraspanin CD9 as novel binding partner for JAM-A in endothelial cells. CD9 acts as scaffold and assembles a ternary JAM-A-CD9-αvβ3 integrin complex from which JAM-A is released upon bFGF stimulation. CD9 interacts predominantly with monomeric JAM-A, which suggests that bFGF induces signaling by triggering JAM-A dimerization. Among the two vitronectin receptors, αvβ3 and αvβ5 integrin, which have been shown to cooperate during angiogenic signaling with bFGF and vascular endothelial growth factor (VEGF), respectively, CD9 links JAM-A specifically to αvβ3 integrin. In line with this, knockdown of CD9 blocks bFGF- but not VEGF-induced ERK1/2 activation. JAM-A or CD9 knockdown impairs endothelial cell migration and tube formation. Our findings indicate that CD9 incorporates monomeric JAM-A into a complex with αvβ3 integrin, which responds to bFGF stimulation by JAM-A release to regulate mitogen-activated protein kinase (MAPK) activation, endothelial cell migration, and angiogenesis. The data also provide new mechanistic insights into the cooperativity between bFGF and αvβ3 integrin during angiogenic signaling.
Figures

References
-
- Arnaoutova I, Kleinman HK. In vitro angiogenesis: endothelial cell tube formation on gelled basement membrane extract. Nature Protoc. 2010;5:628–635. - PubMed
-
- Barreiro O, Yanez-Mo M, Sala-Valdes M, Gutierrez-Lopez MD, Ovalle S, Higginbottom A, Monk PN, Cabanas C, Sanchez-Madrid F. Endothelial tetraspanin microdomains regulate leukocyte firm adhesion during extravasation. Blood. 2005;105:2852–2861. - PubMed
-
- Bazzoni G. The JAM family of junctional adhesion molecules. Curr Opin Cell Biol. 2003;15:525–530. - PubMed
-
- Bazzoni G, Martinez-Estrada OM, Orsenigo F, Cordenonsi M, Citi S, Dejana E. Interaction of junctional adhesion molecule with the tight junction components ZO-1, cingulin, and occludin. J Biol Chem. 2000;275:20520–20526. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
