

Critical residues in the PMEL/Pmel17 N-terminus direct the hierarchical assembly of melanosomal fibrils
- PMID: 23389629
- PMCID: PMC3608505
- DOI: 10.1091/mbc.E12-10-0742
Critical residues in the PMEL/Pmel17 N-terminus direct the hierarchical assembly of melanosomal fibrils
Abstract
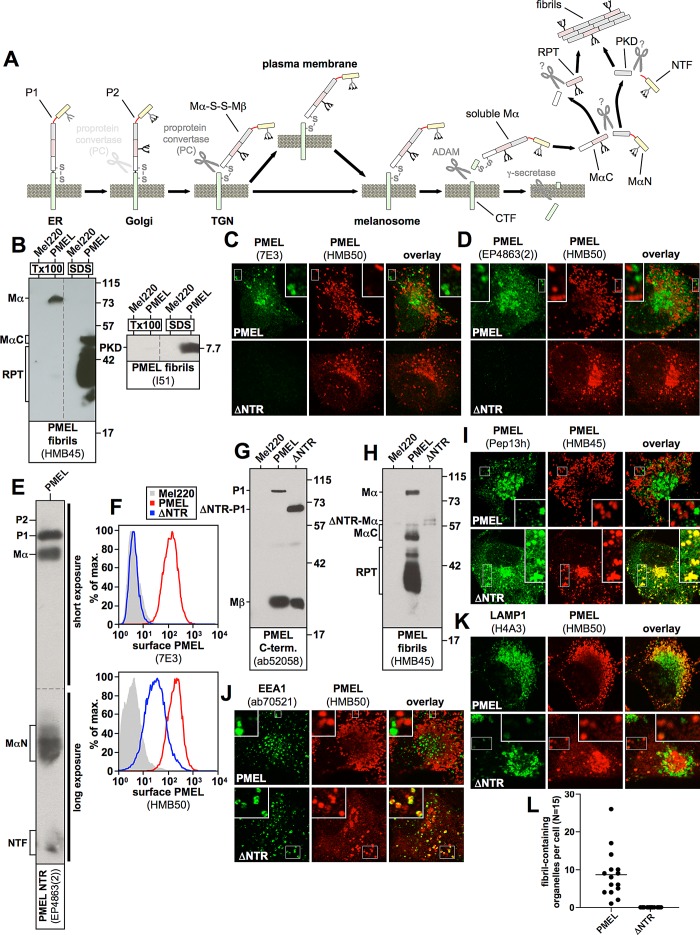
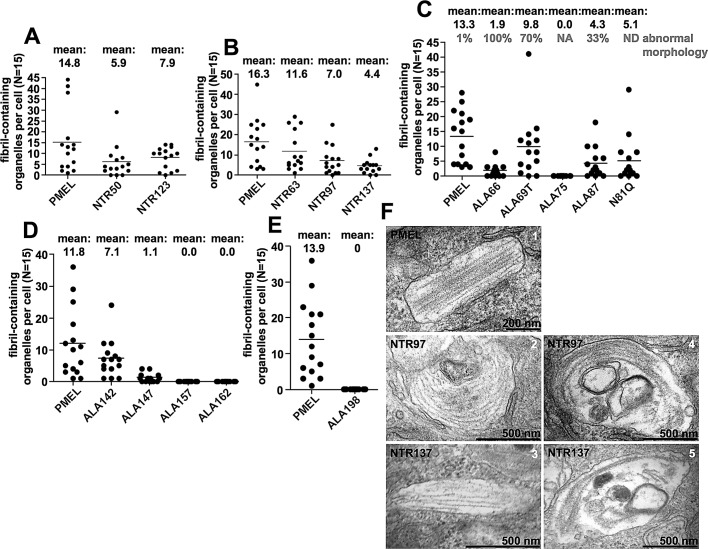
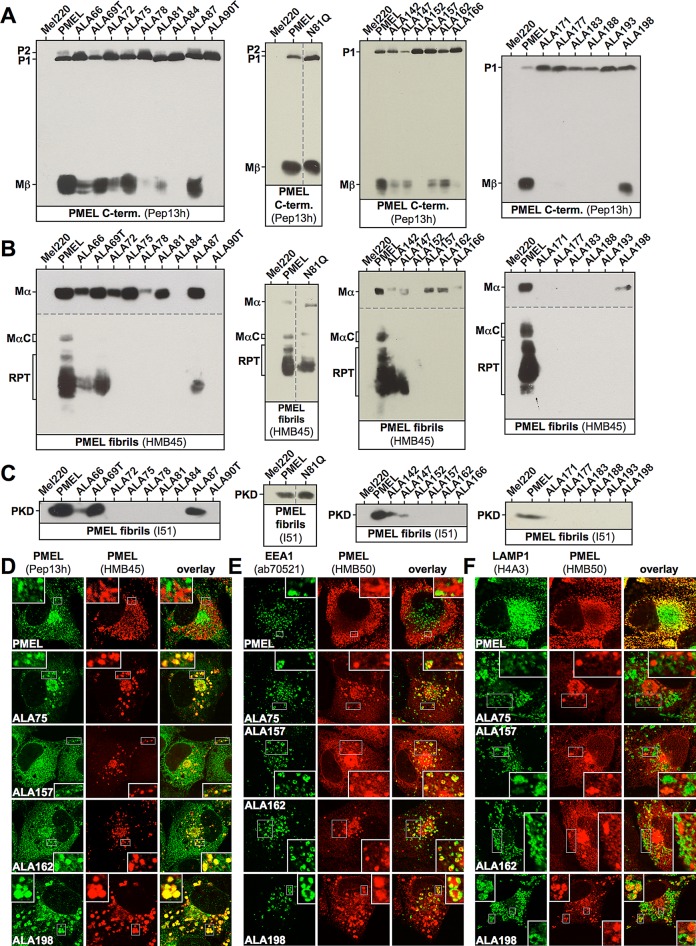
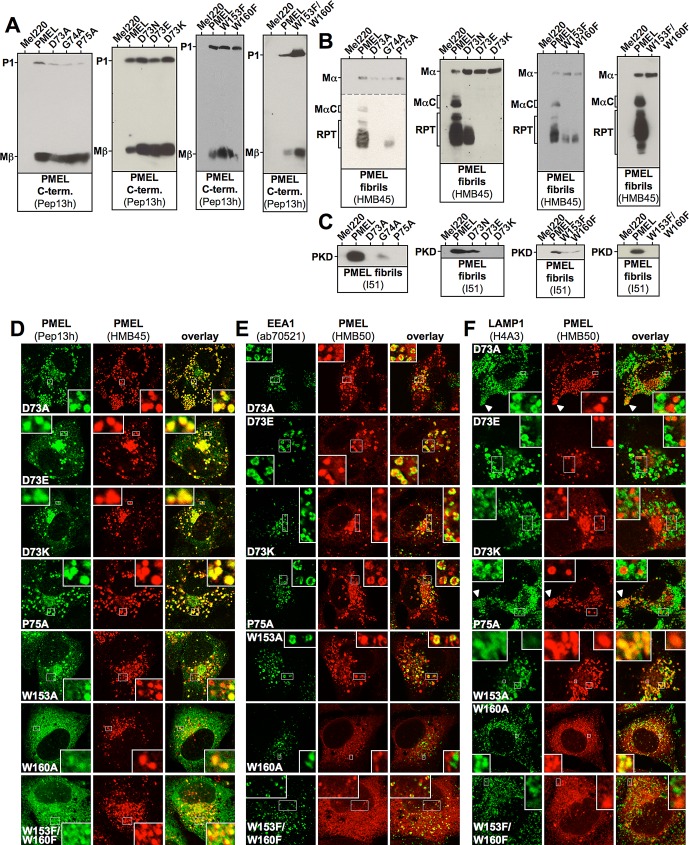
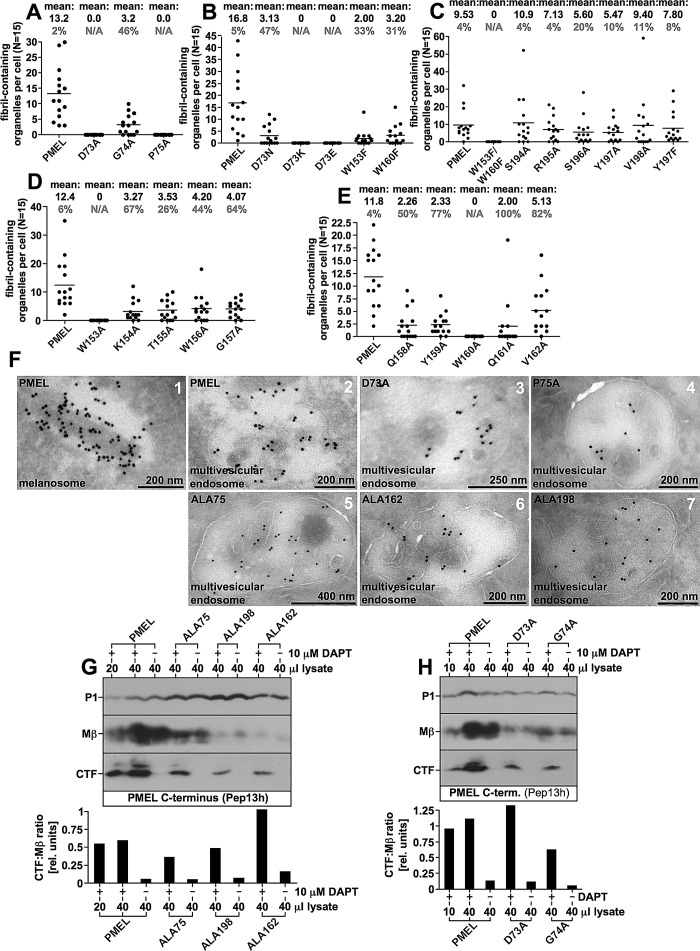
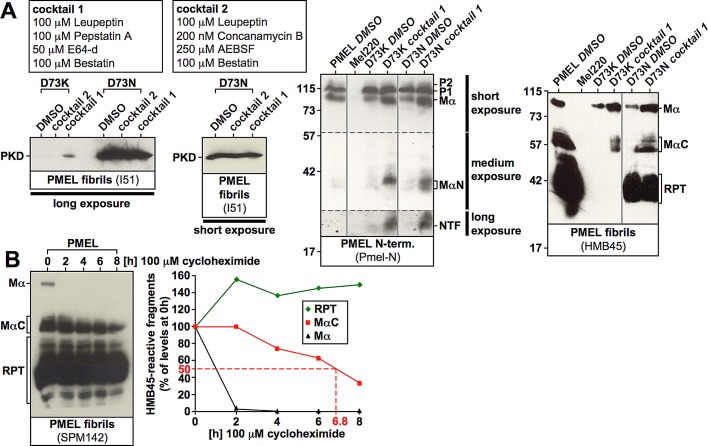
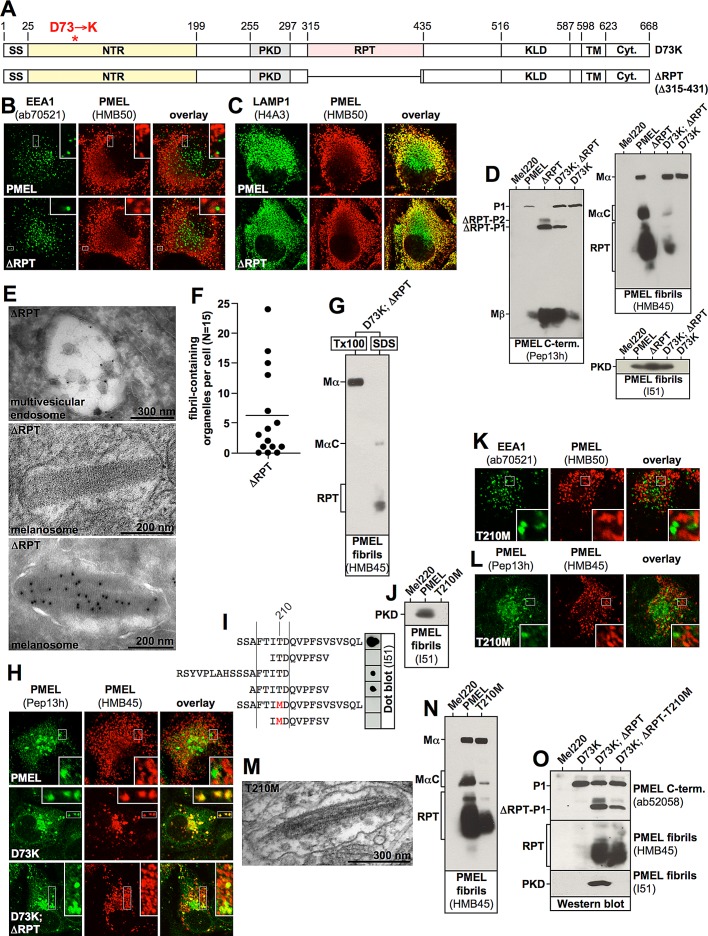
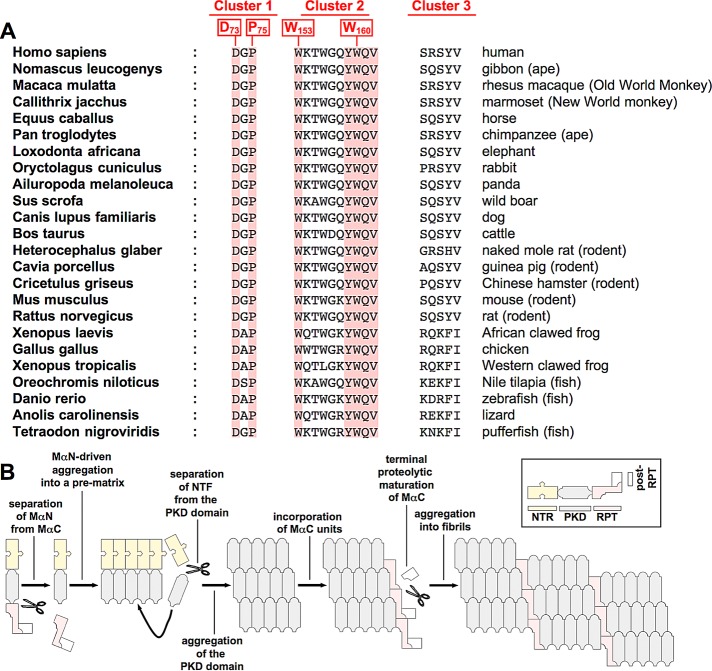
PMEL (also called Pmel17 or gp100) is a melanocyte/melanoma-specific glycoprotein that plays a critical role in melanosome development by forming a fibrillar amyloid matrix in the organelle for melanin deposition. Although ultimately not a component of mature fibrils, the PMEL N-terminal region (NTR) is essential for their formation. By mutational analysis we establish a high-resolution map of this domain in which sequence elements and functionally critical residues are assigned. We show that the NTR functions in cis to drive the aggregation of the downstream polycystic kidney disease (PKD) domain into a melanosomal core matrix. This is essential to promote in trans the stabilization and terminal proteolytic maturation of the repeat (RPT) domain-containing MαC units, precursors of the second fibrillogenic fragment. We conclude that during melanosome biogenesis the NTR controls the hierarchical assembly of melanosomal fibrils.
Figures

References
-
- Bogan AA, Thorn KS. Anatomy of hot spots in protein interfaces. J Mol Biol. 1998;280:1–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
