

Fusing catalase to an alkane-producing enzyme maintains enzymatic activity by converting the inhibitory byproduct H2O2 to the cosubstrate O2
- PMID: 23391732
- PMCID: PMC3581945
- DOI: 10.1073/pnas.1218769110
Fusing catalase to an alkane-producing enzyme maintains enzymatic activity by converting the inhibitory byproduct H2O2 to the cosubstrate O2
Abstract
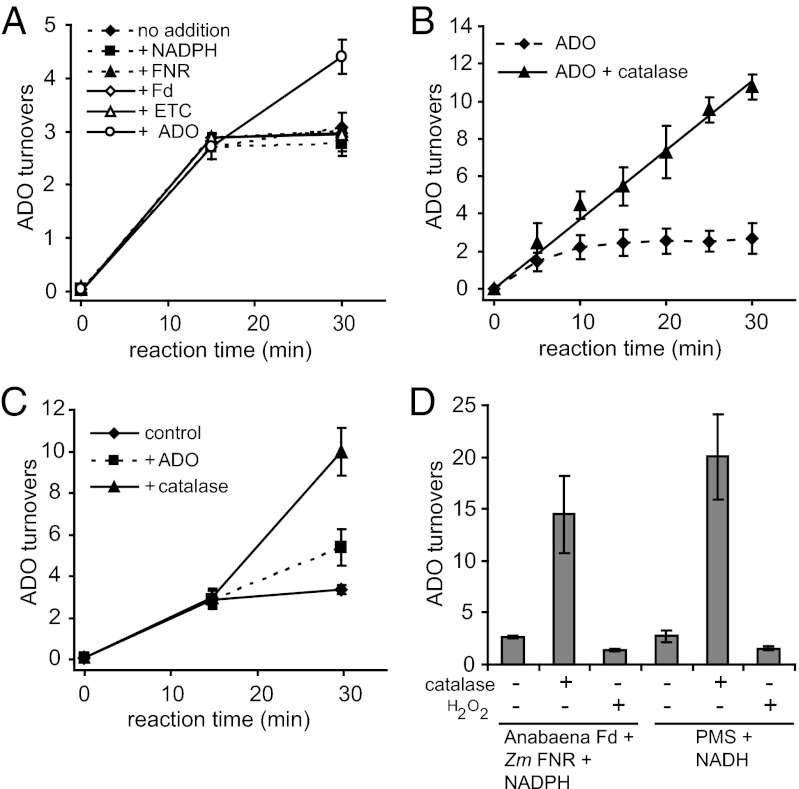
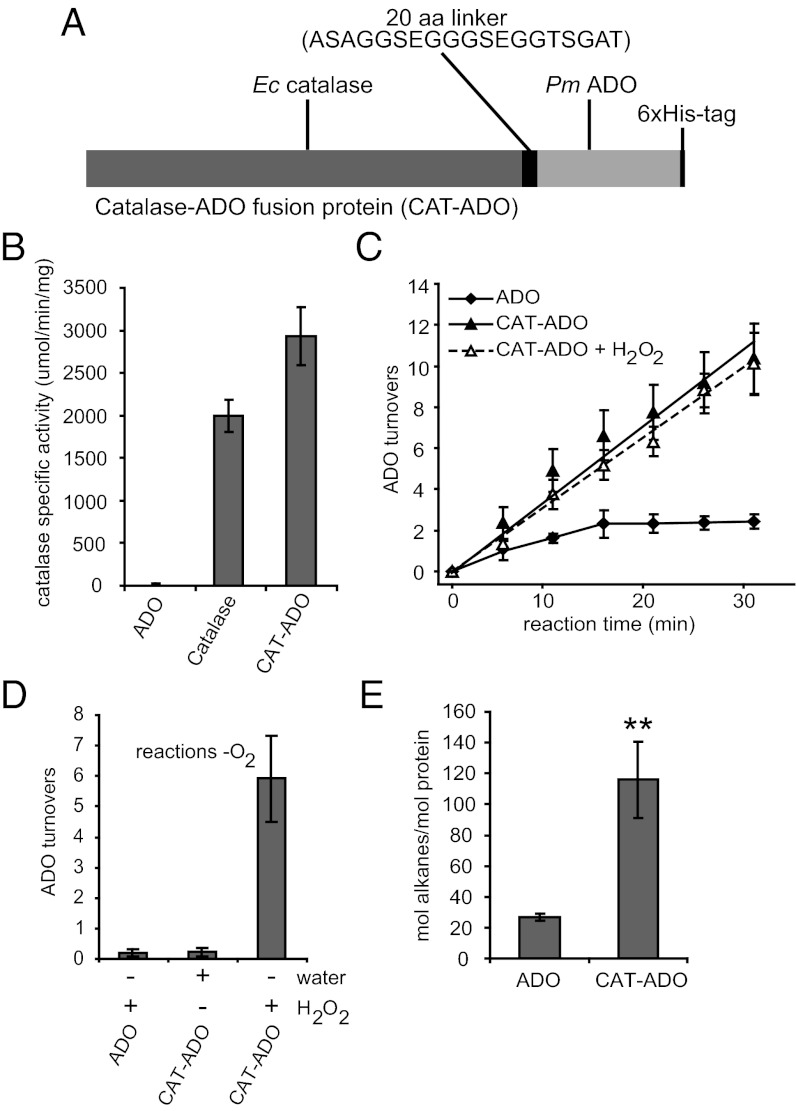
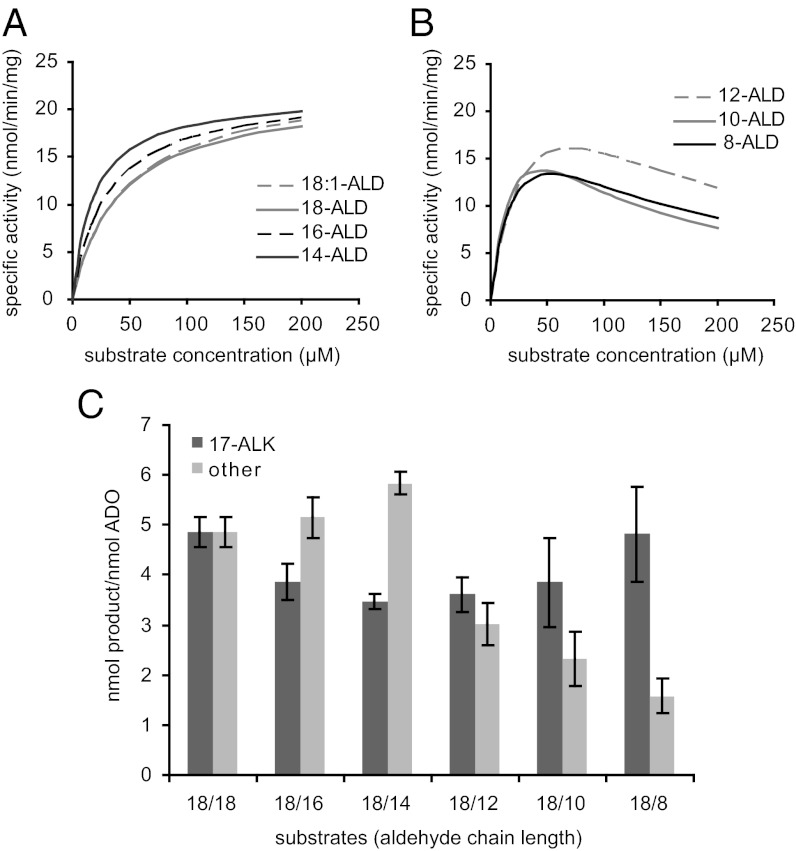
Biologically produced alkanes represent potential renewable alternatives to petroleum-derived chemicals. A cyanobacterial pathway consisting of acyl-Acyl Carrier Protein reductase and an aldehyde-deformylating oxygenase (ADO) converts acyl-Acyl Carrier Proteins into corresponding n-1 alkanes via aldehyde intermediates in an oxygen-dependent manner (K(m) for O(2), 84 ± 9 µM). In vitro, ADO turned over only three times, but addition of more ADO to exhausted assays resulted in additional product formation. While evaluating the peroxide shunt to drive ADO catalysis, we discovered that ADO is inhibited by hydrogen peroxide (H(2)O(2)) with an apparent K(i) of 16 ± 6 µM and that H(2)O(2) inhibition is of mixed-type with respect to O(2). Supplementing exhausted assays with catalase (CAT) restored ADO activity, demonstrating that inhibition was reversible and dependent on H(2)O(2), which originated from poor coupling of reductant consumption with alkane formation. Kinetic analysis showed that long-chain (C14-C18) substrates follow Michaelis-Menten kinetics, whereas short and medium chains (C8-C12) exhibit substrate inhibition. A bifunctional protein comprising an N-terminal CAT coupled to a C-terminal ADO (CAT-ADO) prevents H(2)O(2) inhibition by converting it to the cosubstrate O(2). Indeed, alkane production by the fusion protein is observed upon addition of H(2)O(2) to an anaerobic reaction mix. In assays, CAT-ADO turns over 225 times versus three times for the native ADO, and its expression in Escherichia coli increases catalytic turnovers per active site by fivefold relative to the expression of native ADO. We propose the term "protection via inhibitor metabolism" for fusion proteins designed to metabolize inhibitors into noninhibitory compounds.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Reed JR, Quilici DR, Blomquist GJ, Reitz RC. Proposed mechanism for the cytochrome P450-catalyzed conversion of aldehydes to hydrocarbons in the house fly, Musca domestica. Biochemistry. 1995;34(49):16221–16227. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
