

Review
doi: 10.1038/nri3386.
Epub 2013 Feb 8.
Circadian control of the immune system
Affiliations
- PMID: 23391992
- PMCID: PMC4090048
- DOI: 10.1038/nri3386
Item in Clipboard
Review
Circadian control of the immune system
Nat Rev Immunol.
2013 Mar.
Abstract
Circadian rhythms, which have long been known to play crucial roles in physiology, are emerging as important regulators of specific immune functions. Circadian oscillations of immune mediators coincide with the activity of the immune system, possibly allowing the host to anticipate and handle microbial threats more efficiently. These oscillations may also help to promote tissue recovery and the clearance of potentially harmful cellular elements from the circulation. This Review summarizes the current knowledge of circadian rhythms in the immune system and provides an outlook on potential future implications.
Figures
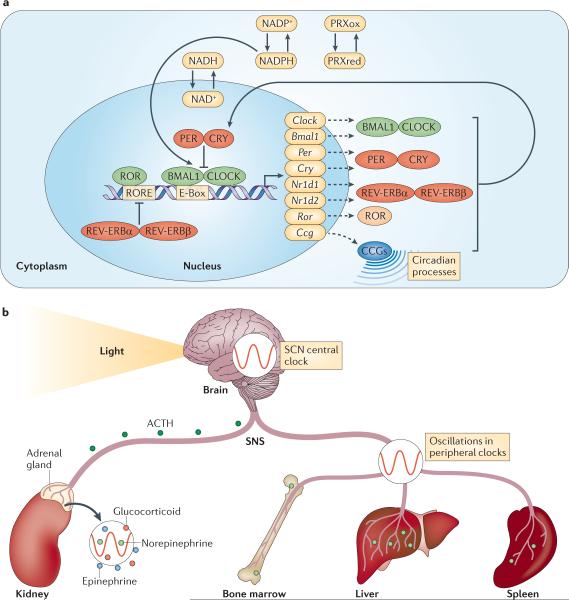
a) The molecular clock. Transcription of the core clock genes Bmal1 and Clock (or its related gene Npas2, not shown) results in their heterodimerization in the cytoplasm and ensuing nuclear translocation. These helix-loop-helix transcription factors bind to canonical E-Box sequences (CACGTG) of clock-controlled genes (CCGs), driving circadian processes and their own expression. In addition, the expression of negative (PER and CRY) as well as positive (retinoic acid-related orphan receptor (ROR)) regulators of this cycle is induced. The PER/CRY complex represses binding of BMAL1–CLOCK to target genes, whereas ROR binding to ROR responsive elements (ROREs) in the Bmal1 promoter induces expression. After a period of time, the PER/CRY complex is degraded and BMAL1–CLOCK activates another transcription cycle. A second autoregulatory feedback loop is induced by transcription of REV-ERBα and REV-ERBβ (encoded by Nr1d1 and Nr1d2). The REV-ERBα–REV-ERBβ complex represses Bmal1 transcription and competes with ROR for binding of ROREs. While this pathway stabilizes the clock it can also directly drive circadian rhythms. The molecular clock does not only depend on transcriptional/translational feedback but is also regulated by rhythms in post-translational modifications of proteins such as oxidation cycles of peroxiredoxins (PRXox(idized)/PRXred(uced)) as well as NADPH and NADH, the latter of which can modulate the binding of the BMAL1/CLOCK complex to DNA directly. b) Entrainment and synchronization. Circadian rhythms are entrained by external cues, of which light is a major contributor. Light is processed via the retina, leading to synchronization of rhythms in hypothalamic suprachiasmatic nuclei, which comprise the master clock of the organism. From here, humoral and neural output systems modulate clocks in peripheral tissues via the hypothalamic-pituitary-adrenal (HPA) axis, setting a common phase. Release of adrenocorticotropic hormone (ACTH) from the pituitary gland likely cooperates with the sympathetic nervous system (SNS) to regulate rhythmic release of hormones (glucocorticoids, epinephrine and norepinephrine) from the adrenal glands. In addition the SNS directly innervates tissues, and can modulate circadian rhythms locally via cyclical release of norepinephrine from nerve varicosities.
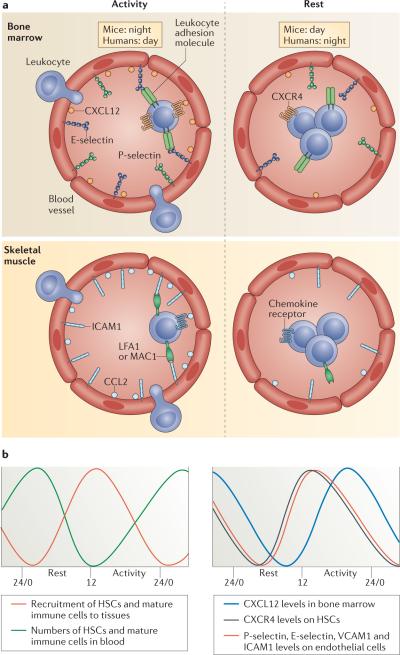
a) Rest-activity cycles in diurnal humans and nocturnal mice. In murine bone marrow and skeletal muscle, leukocyte recruitment is enhanced during the active phase due to higher expression levels of chemokines as well as adhesion molecules on both the endothelium and hematopoietic cells. In peripheral tissues, the molecules oscillating on leukocytes mediating circadian recruitment are not known but likely include CCR2, the receptor for CCL2, and the ICAM-1 binding integrins LFA-1 and Mac-1. During the resting phase, lower adhesion molecule and chemokine expression as well as enhanced mobilization from the bone marrow results in reduced leukocyte recruitment and enhanced numbers of immune cells in blood. b) Oscillation of factors known to contribute to circadian trafficking of hematopoietic cells showing in-phase expression of pro-migratory factors.
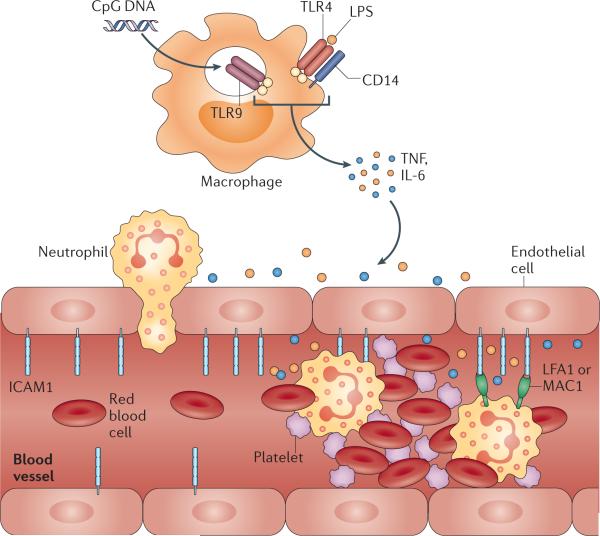
Cyclical exposure to pathogens or other inflammatory insults is matched by oscillations in the expression of pattern recognition receptors (PPRs) on tissue macrophages such as toll-like receptor 9 (TLR9) and TLR4-associated factors. These recognize microbial CpG DNA and lipopolysaccharides (LPS), respectively, which triggers signaling events that lead to cyclical release of the pro-inflammatory cytokines TNF and IL-6. These cytokines can induce inflammation both locally and systemically, resulting in the recruitment of leukocytes from the circulation to combat the infection. Leukocyte trafficking is under circadian control, with circadian release from the bone marrow and cyclical recruitment to tissues, the latter being due to enhanced expression of pro-migratory factors such as intercellular cell adhesion molecule 1 (ICAM1) on endothelial cells. Rhythmic leukocyte/endothelial cell interactions can contribute to obstructing blood flow in small caliber vessels by exacerbating leukocyte activation, prompting their interactions with other free-flowing blood components such as red blood cells (RBCs), and potentially causing thrombus formation and vascular infarction as observed in sickle cell disease.
References
-
- Halberg F, Halberg E, Barnum CP, Bittner JJ. Physiological 24-hour Periodicity in Human Beings and Mice, the Lighting Regimen and Daily Routine. In: Withrow RB, editor. Photoperiodism and Related Phenomena in Plants and Animals. 1959. pp. 803–78.
-
- Panda S, et al. Coordinated transcription of key pathways in the mouse by the circadian clock. Cell. 2002;109:307–20. - PubMed
-
- Storch KF, et al. Extensive and divergent circadian gene expression in liver and heart. Nature. 2002;417:78–83. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
