

HRG1 is essential for heme transport from the phagolysosome of macrophages during erythrophagocytosis
- PMID: 23395172
- PMCID: PMC3582031
- DOI: 10.1016/j.cmet.2013.01.005
HRG1 is essential for heme transport from the phagolysosome of macrophages during erythrophagocytosis
Abstract
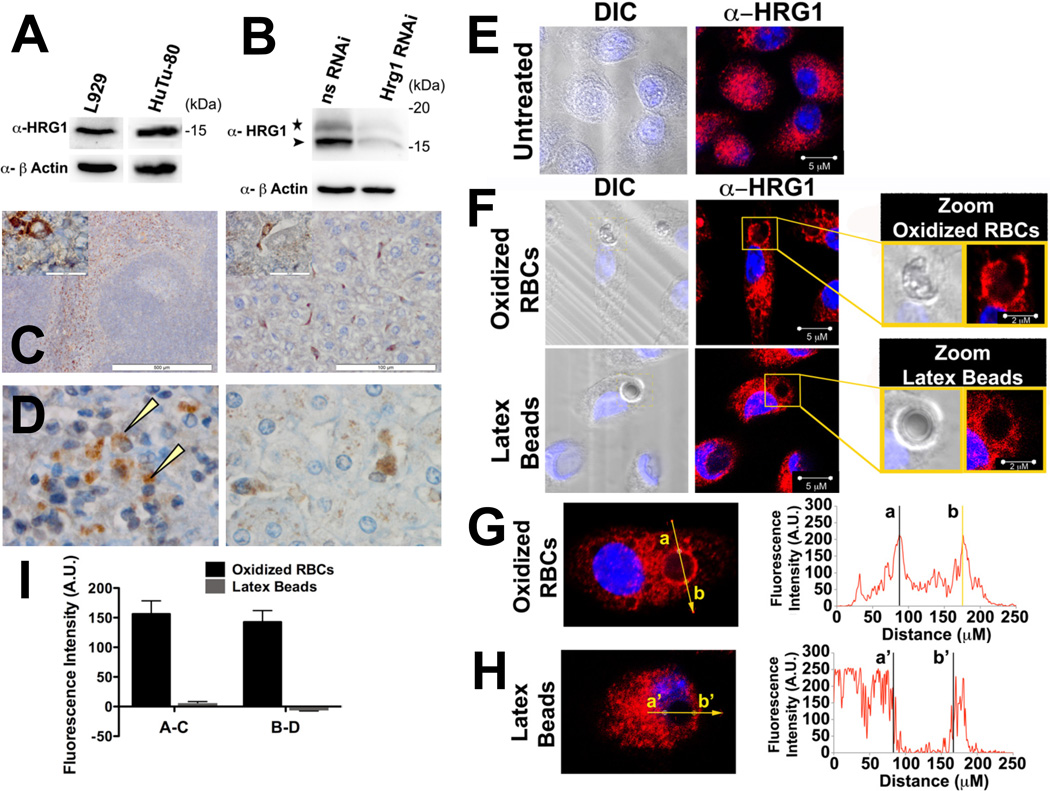
Adult humans have about 25 trillion red blood cells (RBCs), and each second we recycle about 5 million RBCs by erythrophagocytosis (EP) in macrophages of the reticuloendothelial system. Despite the central role for EP in mammalian iron metabolism, the molecules and pathways responsible for heme trafficking during EP remain unknown. Here, we show that the mammalian homolog of HRG1, a transmembrane heme permease in C. elegans, is essential for macrophage iron homeostasis and transports heme from the phagolysosome to the cytoplasm during EP. HRG1 is strongly expressed in macrophages of the reticuloendothelial system and specifically localizes to the phagolysosomal membranes during EP. Depletion of Hrg1 in mouse macrophages causes attenuation of heme transport from the phagolysosomal compartment. Importantly, missense polymorphisms in human HRG1 are defective in heme transport. Our results reveal HRG1 as the long-sought heme transporter for heme-iron recycling in macrophages and suggest that genetic variations in HRG1 could be modifiers of human iron metabolism.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures




Similar articles
-
Hemozoin produced by mammals confers heme tolerance.Elife. 2019 Oct 1;8:e49503. doi: 10.7554/eLife.49503. Elife. 2019. PMID: 31571584 Free PMC article.
-
Hrg1 promotes heme-iron recycling during hemolysis in the zebrafish kidney.PLoS Genet. 2018 Sep 24;14(9):e1007665. doi: 10.1371/journal.pgen.1007665. eCollection 2018 Sep. PLoS Genet. 2018. PMID: 30248094 Free PMC article.
-
Subcellular localization of iron and heme metabolism related proteins at early stages of erythrophagocytosis.PLoS One. 2012;7(7):e42199. doi: 10.1371/journal.pone.0042199. Epub 2012 Jul 30. PLoS One. 2012. PMID: 22860081 Free PMC article.
-
Macrophages and iron trafficking at the birth and death of red cells.Blood. 2015 May 7;125(19):2893-7. doi: 10.1182/blood-2014-12-567776. Epub 2015 Mar 16. Blood. 2015. PMID: 25778532 Free PMC article. Review.
-
[Erythrophagocytosis and recycling of heme iron in normal and pathological conditions; regulation by hepcidin].Transfus Clin Biol. 2005 Jun;12(2):123-30. doi: 10.1016/j.tracli.2005.04.017. Transfus Clin Biol. 2005. PMID: 15927501 Review. French.
Cited by
-
Overdosing on iron: Elevated iron and degenerative brain disorders.Exp Biol Med (Maywood). 2020 Oct;245(16):1444-1473. doi: 10.1177/1535370220953065. Epub 2020 Sep 2. Exp Biol Med (Maywood). 2020. PMID: 32878460 Free PMC article. Review.
-
Research progress on ferroptosis in gliomas (Review).Oncol Lett. 2023 Nov 27;27(1):36. doi: 10.3892/ol.2023.14169. eCollection 2024 Jan. Oncol Lett. 2023. PMID: 38108075 Free PMC article. Review.
-
Iron and Heme Metabolism at the Leishmania-Host Interface.Trends Parasitol. 2020 Mar;36(3):279-289. doi: 10.1016/j.pt.2019.12.010. Epub 2020 Jan 28. Trends Parasitol. 2020. PMID: 32005611 Free PMC article. Review.
-
The Macrophage Iron Signature in Health and Disease.Int J Mol Sci. 2021 Aug 6;22(16):8457. doi: 10.3390/ijms22168457. Int J Mol Sci. 2021. PMID: 34445160 Free PMC article. Review.
-
"Pumping iron"-how macrophages handle iron at the systemic, microenvironmental, and cellular levels.Pflugers Arch. 2017 Apr;469(3-4):397-418. doi: 10.1007/s00424-017-1944-8. Epub 2017 Mar 1. Pflugers Arch. 2017. PMID: 28251312 Free PMC article. Review.
References
-
- Abboud S, Haile DJ. A novel mammalian iron-regulated protein involved in intracellular iron metabolism. J Biol Chem. 2000;275:19906–19912. - PubMed
-
- Adams PC, Reboussin DM, Barton JC, McLaren CE, Eckfeldt JH, McLaren GD, Dawkins FW, Acton RT, Harris EL, Gordeuk VR, et al. Hemochromatosis and iron-overload screening in a racially diverse population. N Engl J Med. 2005;352:1769–1778. - PubMed
-
- Andrews NC. Disorders of iron metabolism. N Engl J Med. 1999a;341:1986–1995. - PubMed
-
- Andrews NC. Medical Progress: Disorders of iron metabolism. The New England journal of medicine. 1999b;341:1986–1995. - PubMed
-
- Bard F, Casano L, Mallabiabarrena A, Wallace E, Saito K, Kitayama H, Guizzunti G, Hu Y, Wendler F, Dasgupta R, et al. Functional genomics reveals genes involved in protein secretion and Golgi organization. Nature. 2006;439:604–607. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
