

Review
doi: 10.1016/j.molcel.2013.01.018.
Understanding metabolic regulation and its influence on cell physiology
Affiliations
- PMID: 23395269
- PMCID: PMC3569837
- DOI: 10.1016/j.molcel.2013.01.018
Item in Clipboard
Review
Understanding metabolic regulation and its influence on cell physiology
Mol Cell.
.
Abstract
Metabolism impacts all cellular functions and plays a fundamental role in biology. In the last century, our knowledge of metabolic pathway architecture and the genomic landscape of disease has increased exponentially. Combined with these insights, advances in analytical methods for quantifying metabolites and systems approaches to analyze these data now provide powerful tools to study metabolic regulation. Here we review the diverse mechanisms cells use to adapt metabolism to specific physiological states and discuss how metabolic flux analyses can be applied to identify important regulatory nodes to understand normal and pathological cell physiology.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
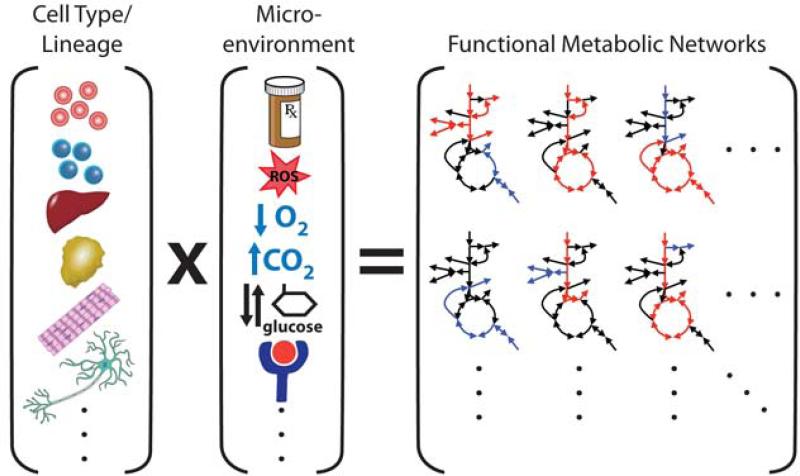
Cells express lineage-specific networks of metabolic enzymes and regulatory factors that support appropriate tissue functions. Microenvironments, hormonal cues, and/or pharmacological perturbations will elicit adaptive metabolic responses that are unique to the metabolic networks of individual cells. Thus, a diverse set of metabolic phenotypes is observed in normal and disease states.
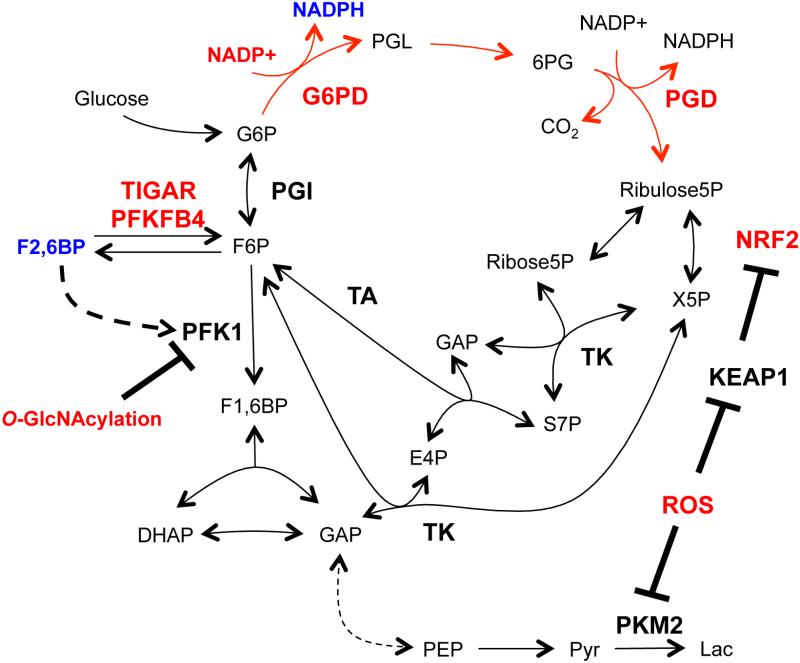
The oxidative pentose phosphate pathway (PPP; red arrows) generates NADPH to maintain reduced glutathione, combat reactive oxygen species (ROS), and maintain redox homeostasis. Transcriptional events and direct enzyme regulation via metabolic intermediates and PTMs mediate both long-lasting and rapid responses to oxidative stress by modulating oxidative PPP flux. Factors that increase oxidative PPP flux are shown in red, while those that reduce flux are shown in blue.
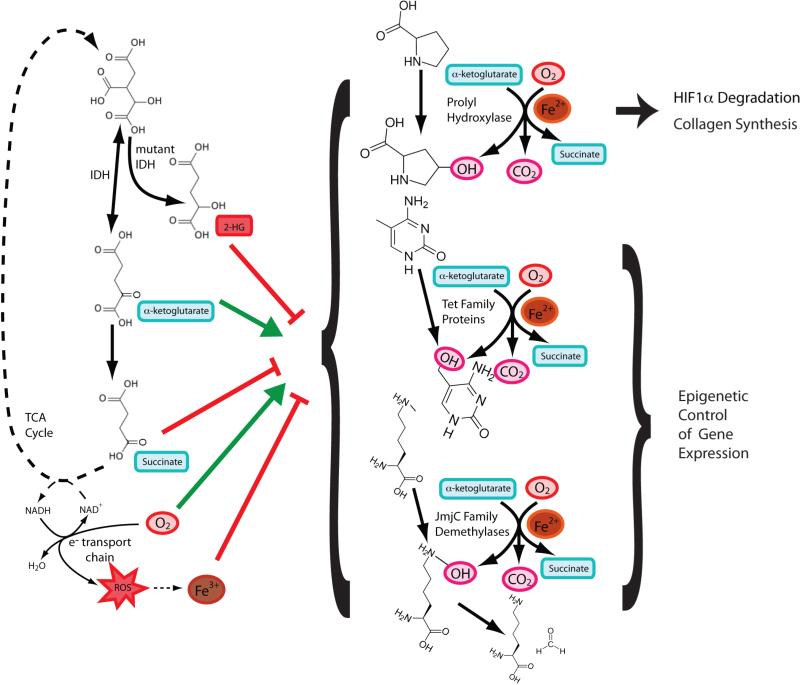
The prolyl hydroxylases, TET proteins, and JmjC domain-containing proteins all carry out similar enzymatic reactions. These dioxygenases split molecular oxygen in an Fe2+-dependent reaction where one oxygen is transferred as a hydroxyl group and the other oxygen is used in a decarboxylation reaction the converts αKG to succinate. Each family of proteins has different hydroxylation substrates. All of these enzymes are influenced by events affecting the redox state of Fe or any of the substrates and products, as well as by levels of (D)-2-hydroxyglutarate, succinate, or fumarate that accumulate as a result of cancer-associated mutations in TCA cycle enzymes.
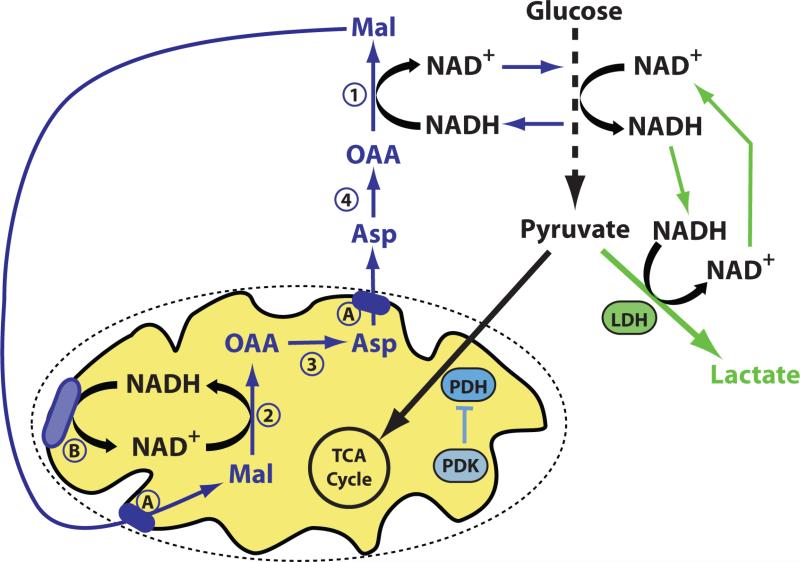
Increased LDH activity and/or decreased PDH activity can shunt glucose-derived pyruvate to lactate. However, metabolism of glucose to pyruvate requires conversion of NAD+ to NADH, and NAD+ must be regenerated from NADH for glycolysis to continue. Pyruvate to lactate conversion efficiently recycles NADH back to NAD+, while oxidation of NADH to NAD+ involving the mitochondria requires four separate enzymatic reactions (numbered 1-4), metabolite transport across the mitochondrial membranes (A), and coupling to the electron transport chain (B) (or other routes of mitochondrial NADH oxidation).
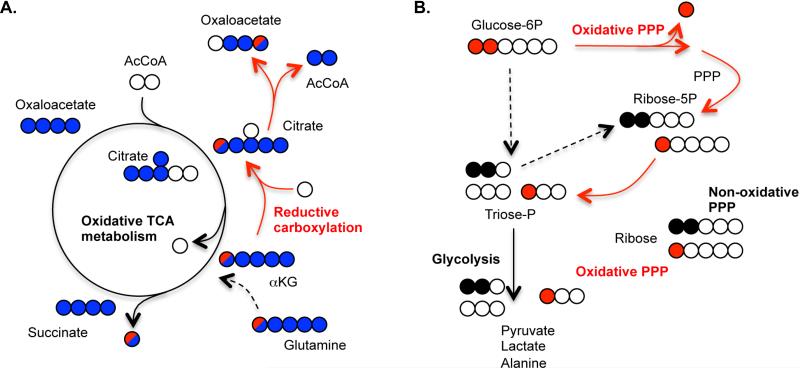
A) Uniformly labeled [U-13C5]glutamine and singly labeled [1-13C]glutamine provide independent means of distinguishing reductive carboxylation from oxidative TCA metabolism when measuring 13C enrichment in various intermediates. Five carbon labeled citrate derived from uniformly labeled glutamine suggests reductive carboxylation while four carbon labeled citrate suggests oxidative TCA metabolism (blue circles). Five carbon labeled citrate can also be obtained from glutamine via pyruvate cycling (not shown), so label incorporation into citrate, oxaloacetate, and downstream metabolites from [1-13C]glutamine provides an independent assessment of reductive TCA flux (red circle). B) Glucose labeled on only the first and second carbons ([1,2-13C2]glucose) is often used to determine relative flux through the oxidative and non-oxidative PPP by assessment of singly versus doubly-labeled downstream intermediates.
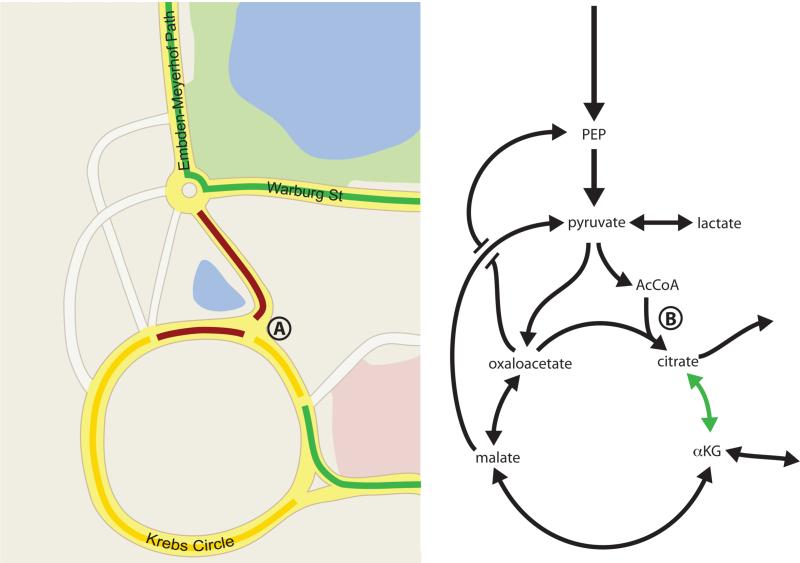
The available routes as well as congestion on each road will affect how quickly cars can navigate between two points. High traffic volume into a point where two roads merge (such as point A on the map) will cause a slowing of traffic as depicted in yellow or red, and result in fewer cars moving via that route per unit time. Road closures or traffic signals can further affect flux though the network of roads as depicted here in the style of Google Maps. Similar principles affect metabolite flow through reaction networks. Decreased flux from AcCoA into the TCA cycle at point B leads to a drop in citrate levels, allowing increased flux from αKG to citrate (green arrow). This example illustrates how a large change in enzyme level may have little effect if metabolite traffic is not constrained at that node, while small changes in enzyme activity can have dramatic effects on flux if a step is limiting. Thus, analysis of individual metabolites, enzyme levels, or maximal enzymatic capacities for single steps may or may not be informative of overall flux.
References
-
- Albe KR, Butler MH, Wright BE. Cellular concentrations of enzymes and their substrates. J Theor Biol. 1990;143:163–195. - PubMed
-
- Antoniewicz MR, Kelleher JK, Stephanopoulos G. Determination of confidence intervals of metabolic fluxes estimated from stable isotope measurements. Metab Eng. 2006;8:324–337. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
