

The ribosome as a hub for protein quality control
- PMID: 23395271
- PMCID: PMC3593112
- DOI: 10.1016/j.molcel.2013.01.020
The ribosome as a hub for protein quality control
Abstract
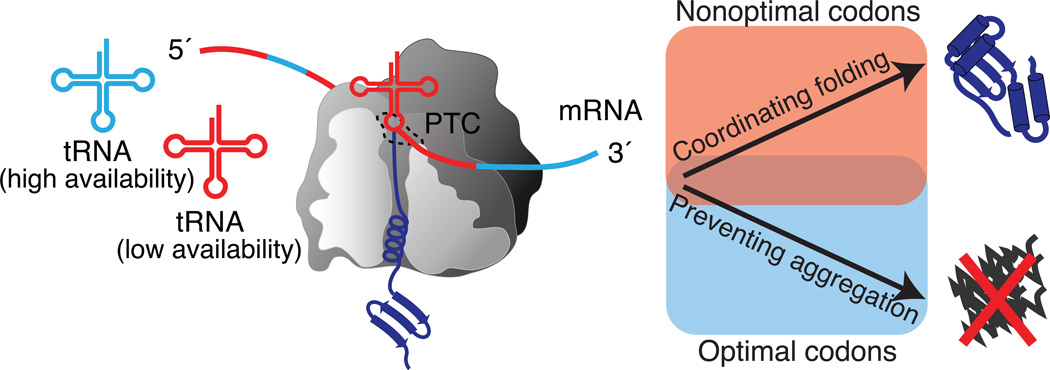
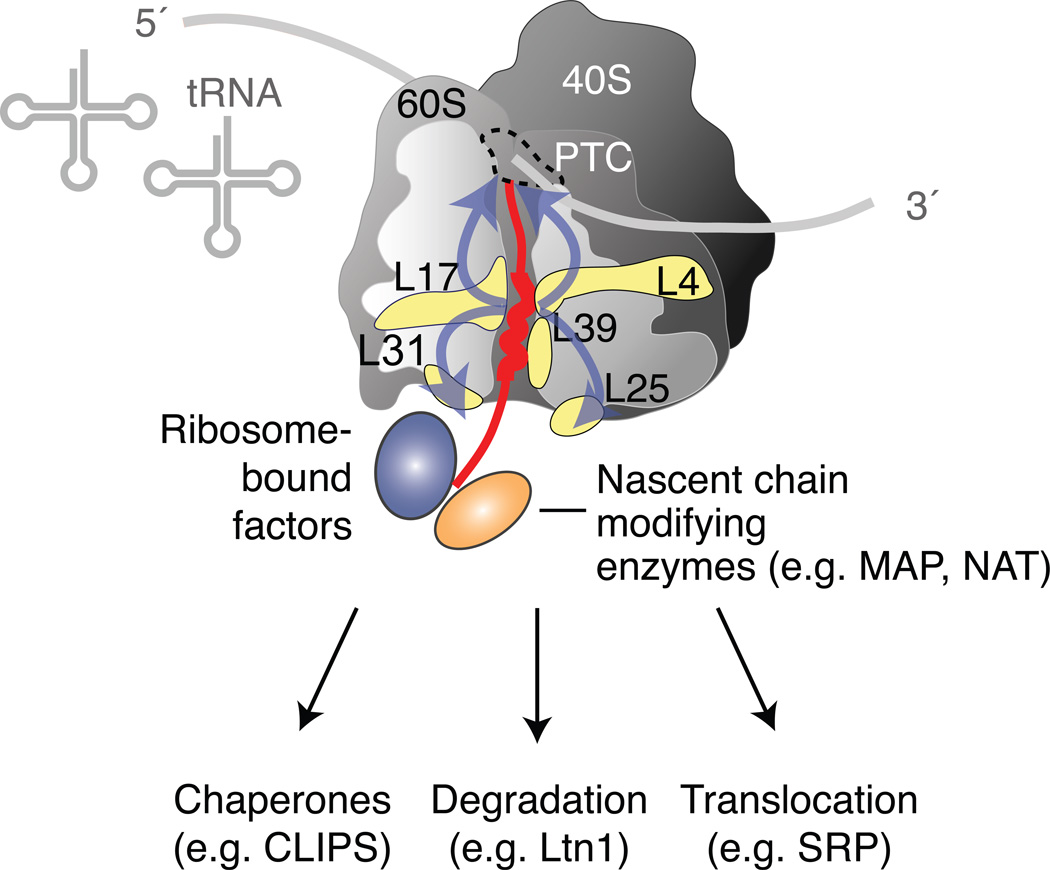
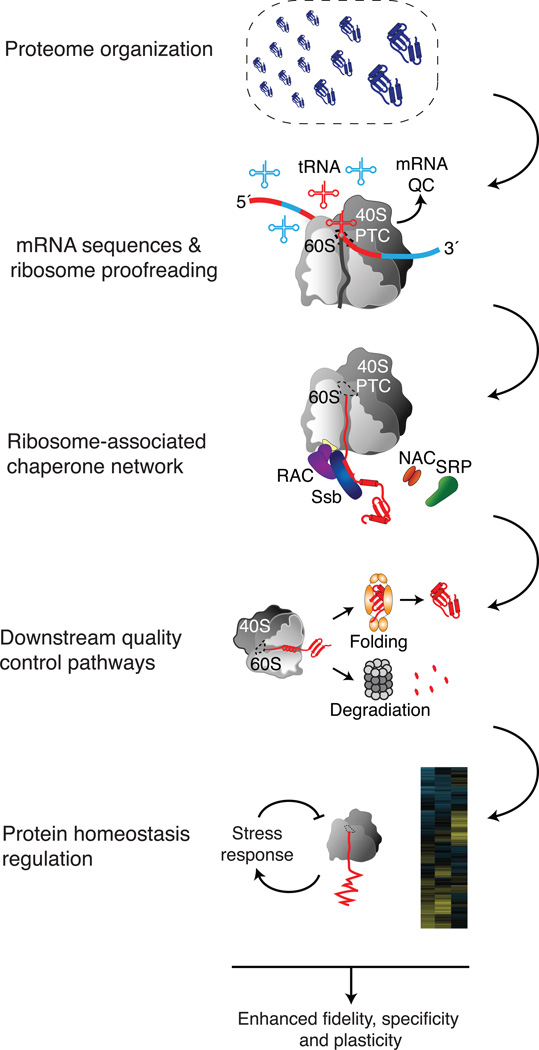
Cells face a constant challenge as they produce new proteins. The newly synthesized polypeptides must be folded properly to avoid aggregation. If proteins do misfold, they must be cleared to maintain a functional and healthy proteome. Recent work is revealing the complex mechanisms that work cotranslationally to ensure protein quality control during biogenesis at the ribosome. Indeed, the ribosome is emerging as a central hub in coordinating these processes, particularly in sensing the nature of the nascent protein chain, recruiting protein folding and translocation components, and integrating mRNA and nascent chain quality control. The tiered and complementary nature of these decision-making processes confers robustness and fidelity to protein homeostasis during protein synthesis.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures

References
-
- Agashe V, Guha S, Chang H-C, Genevaux P, Hayer-Hartl M, Stemp M, Georgopoulos C, Hartl F, Barral J. Function of trigger factor and DnaK in multidomain protein folding: increase in yield at the expense of folding speed. Cell. 2004;117:199–209. - PubMed
-
- Albanèse V, Yam AY-W, Baughman J, Parnot C, Frydman J. Systems analyses reveal two chaperone networks with distinct functions in eukaryotic cells. Cell. 2006;124:75–88. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
