

miR-126 and miR-126* repress recruitment of mesenchymal stem cells and inflammatory monocytes to inhibit breast cancer metastasis
- PMID: 23396050
- PMCID: PMC3672398
- DOI: 10.1038/ncb2690
miR-126 and miR-126* repress recruitment of mesenchymal stem cells and inflammatory monocytes to inhibit breast cancer metastasis
Abstract
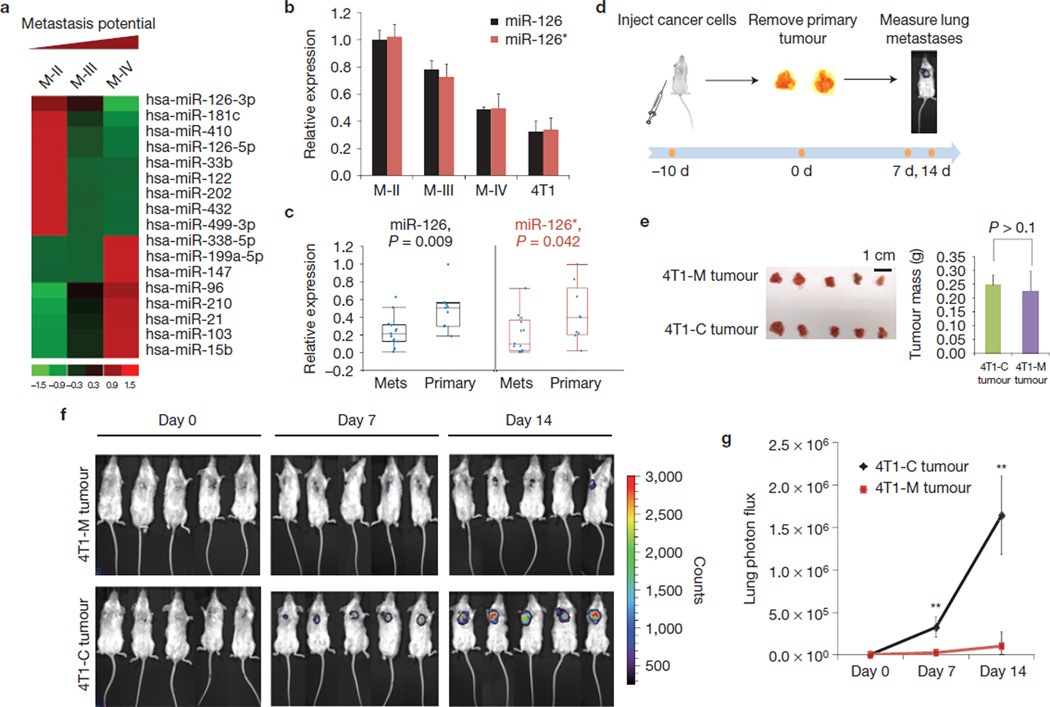
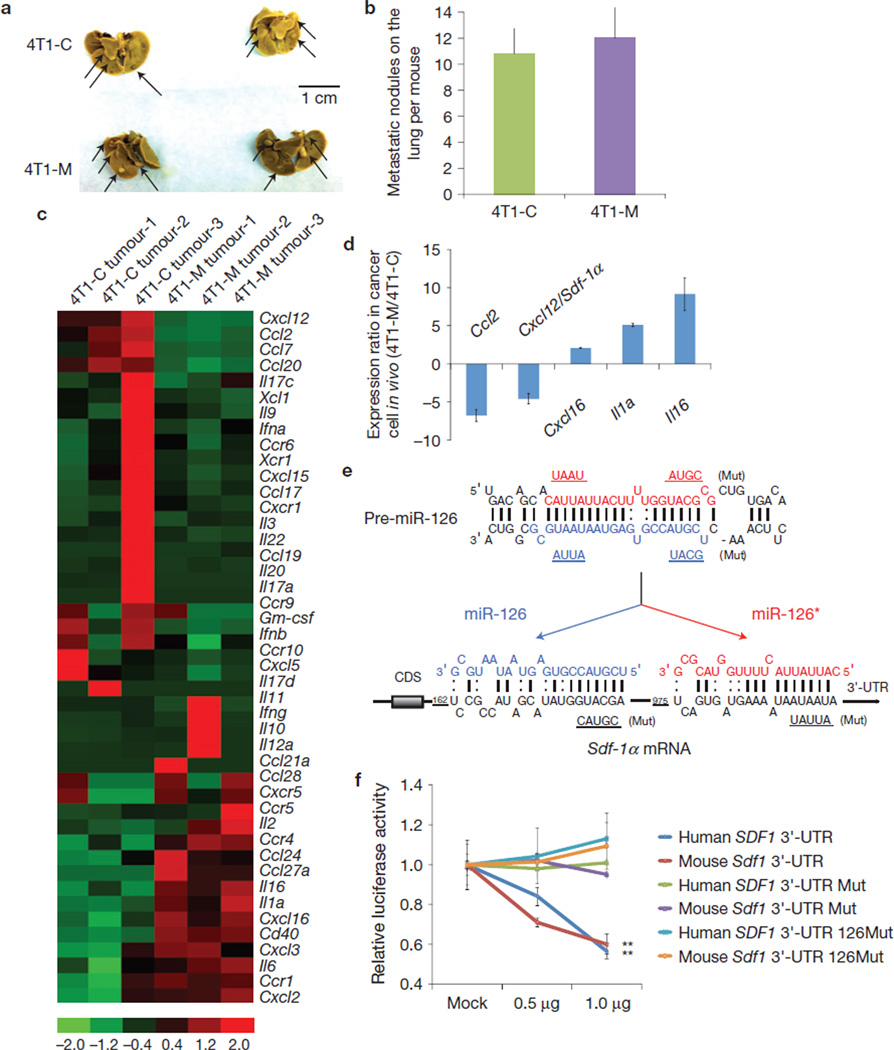
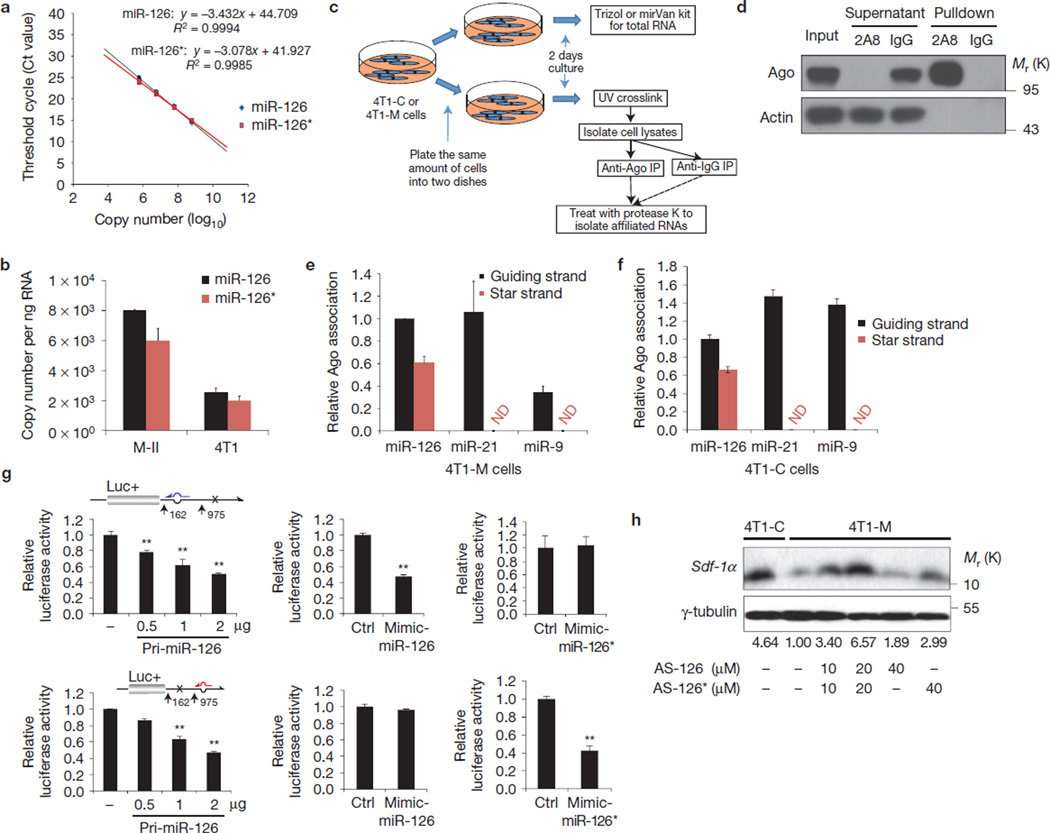
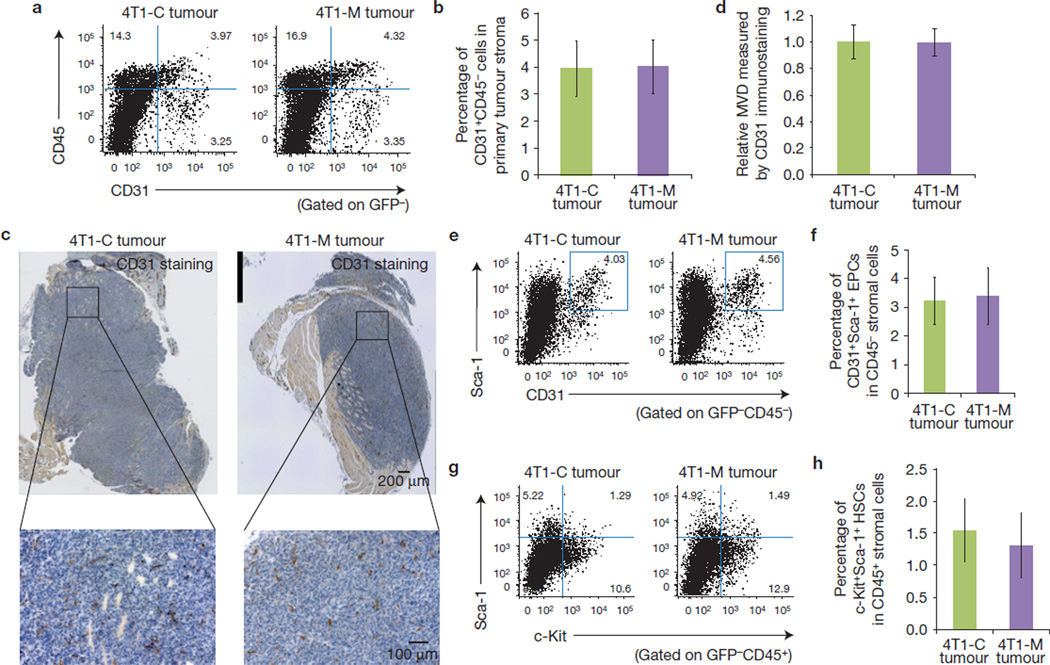
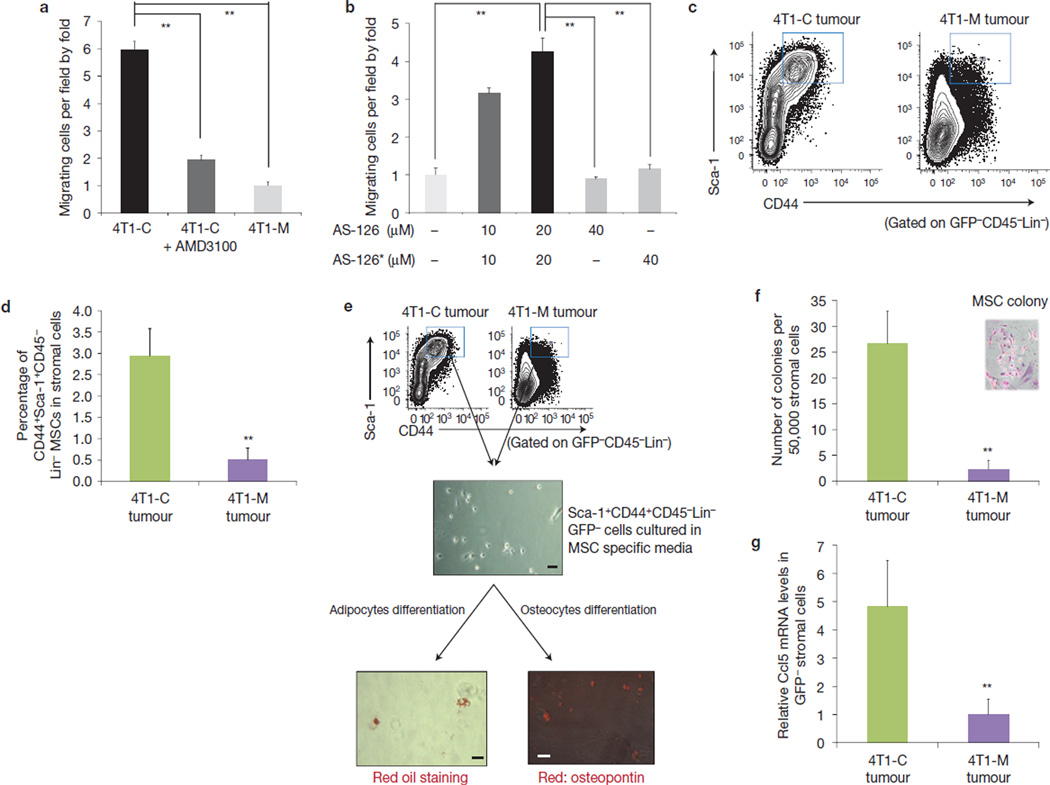
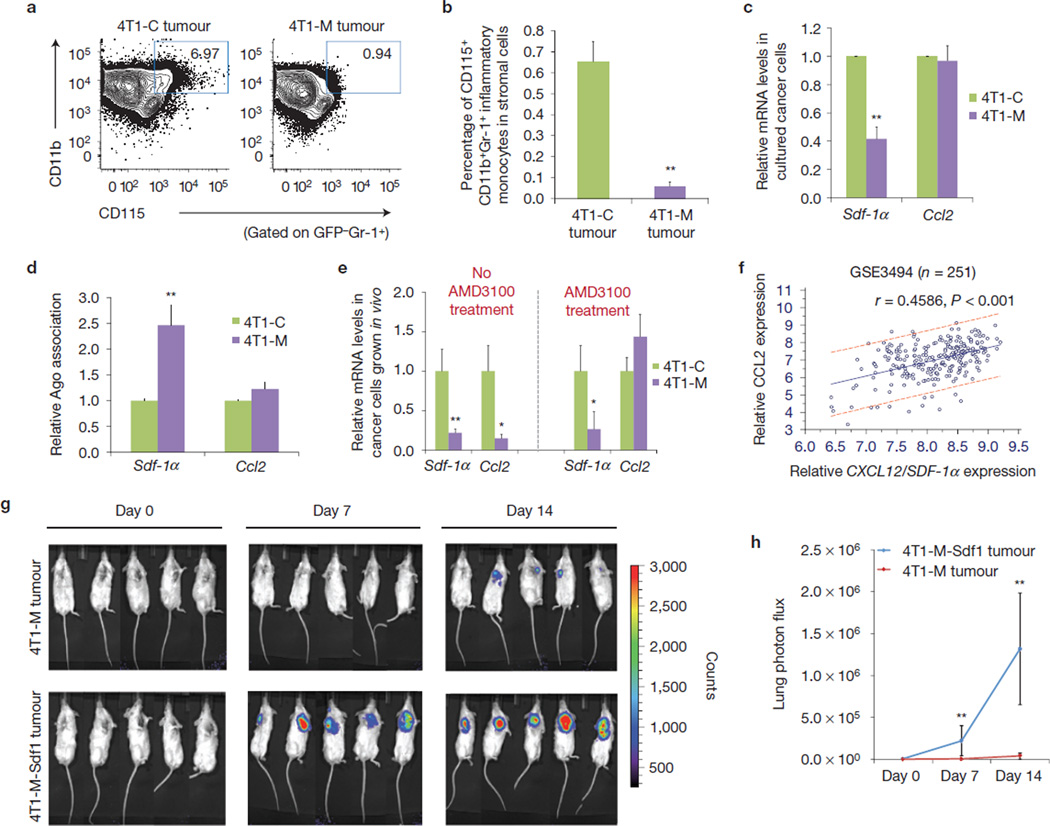
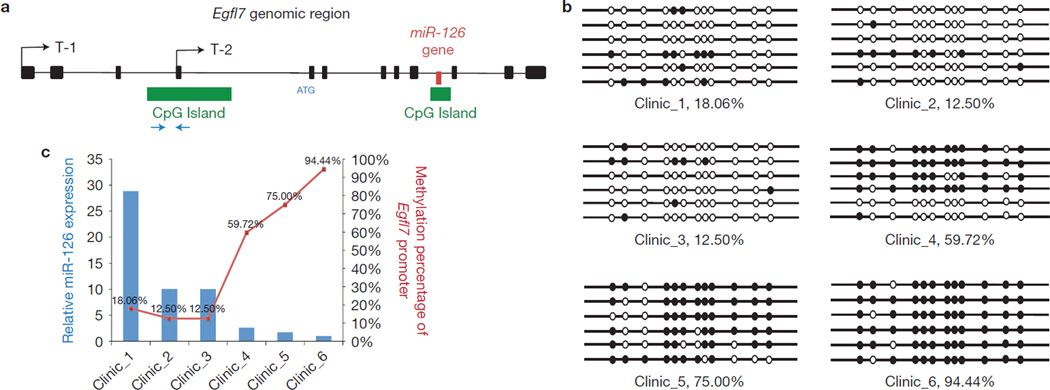
The tumour stroma is an active participant during cancer progression. Stromal cells promote tumour progression and metastasis through multiple mechanisms including enhancing tumour invasiveness and angiogenesis, and suppressing immune surveillance. We report here that miR-126/miR-126(*), a microRNA pair derived from a single precursor, independently suppress the sequential recruitment of mesenchymal stem cells and inflammatory monocytes into the tumour stroma to inhibit lung metastasis by breast tumour cells in a mouse xenograft model. miR-126/miR-126(*) directly inhibit stromal cell-derived factor-1 alpha (SDF-1α) expression, and indirectly suppress the expression of chemokine (C-C motif) ligand 2 (Ccl2) by cancer cells in an SDF-1α-dependent manner. miR-126/miR-126(*) expression is downregulated in cancer cells by promoter methylation of their host gene Egfl7. These findings determine how this microRNA pair alters the composition of the primary tumour microenvironment to favour breast cancer metastasis, and demonstrate a correlation between miR-126/126(*) downregulation and poor metastasis-free survival of breast cancer patients.
Figures
Comment in
-
A one-two punch of miR-126/126* against metastasis.Nat Cell Biol. 2013 Mar;15(3):231-3. doi: 10.1038/ncb2703. Nat Cell Biol. 2013. PMID: 23449143
References
-
- Parkin DM, Bray F, Ferlay J, Pisani P. Global cancer statistics, 2002. CA Cancer J. Clin. 2005;55:74–108. - PubMed
-
- Nguyen DX, Bos PD, Massague J. Metastasis: from dissemination to organ-specific colonization. Nat. Rev. Cancer. 2009;9:274–284. - PubMed
-
- Chaffer CL, Weinberg RA. A perspective on cancer cell metastasis. Science. 2011;331:1559–1564. - PubMed
-
- Hanahan D, Coussens LM. Accessories to the crime: functions of cells recruited to the tumour microenvironment. Cancer Cell. 2012;21:309–322. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
