

Transcriptome profiling and genome-wide DNA binding define the differential role of fenretinide and all-trans RA in regulating the death and survival of human hepatocellular carcinoma Huh7 cells
- PMID: 23396089
- PMCID: PMC3857153
- DOI: 10.1016/j.bcp.2013.01.023
Transcriptome profiling and genome-wide DNA binding define the differential role of fenretinide and all-trans RA in regulating the death and survival of human hepatocellular carcinoma Huh7 cells
Abstract
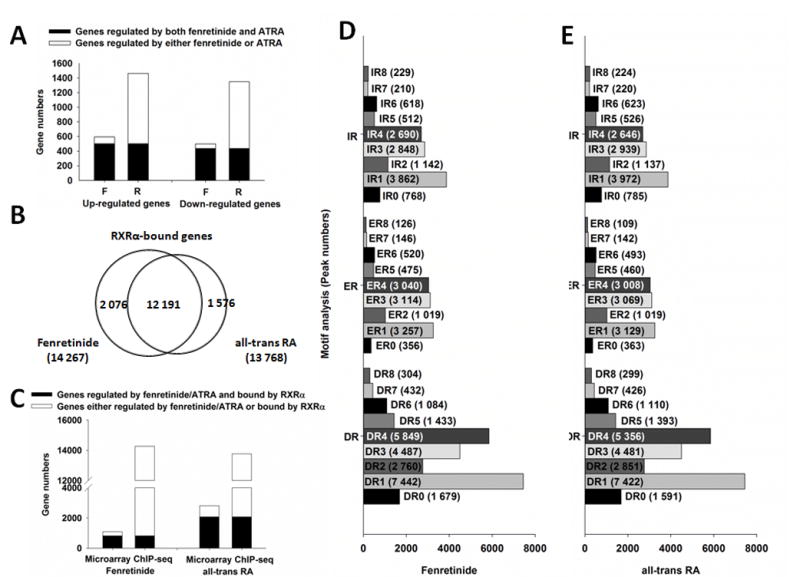
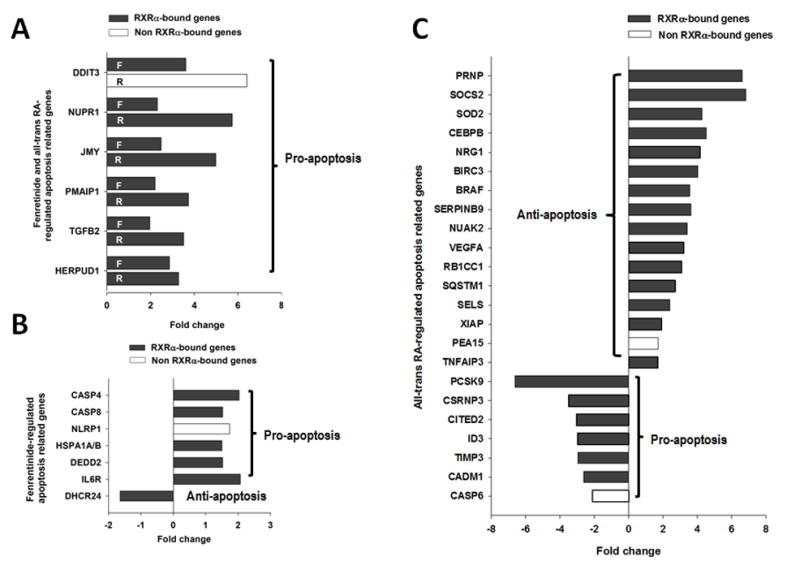
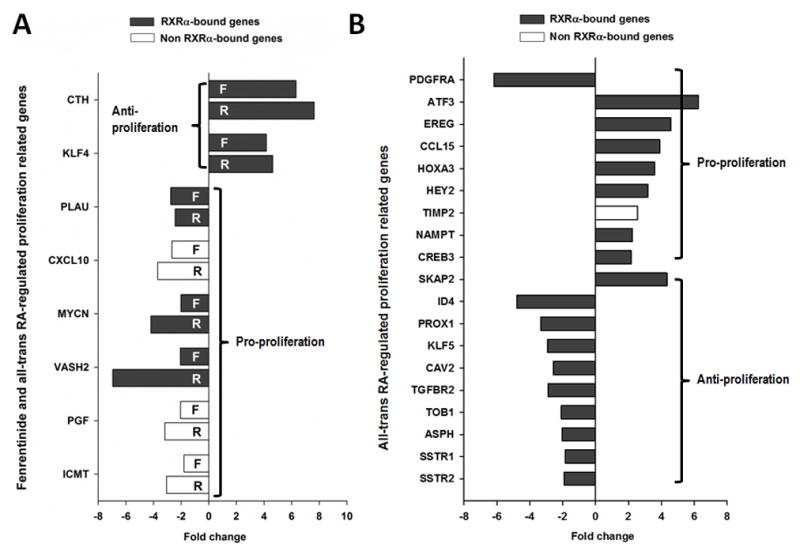
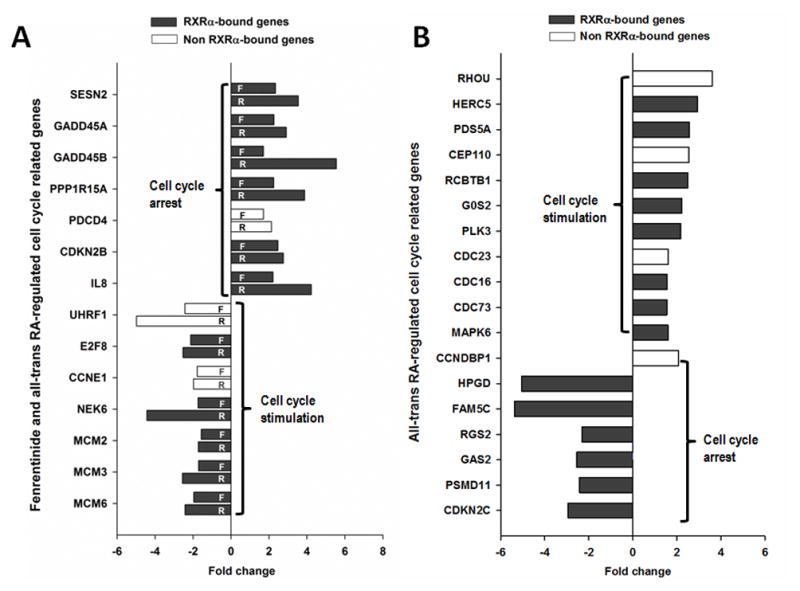
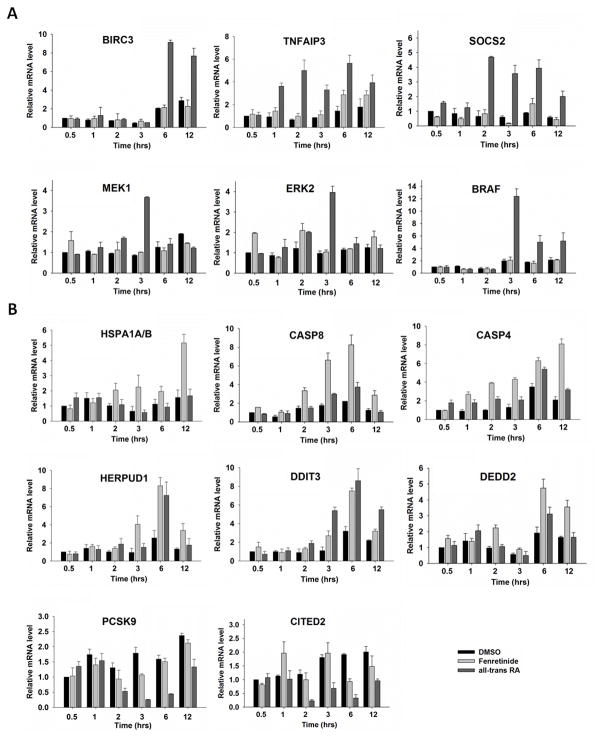
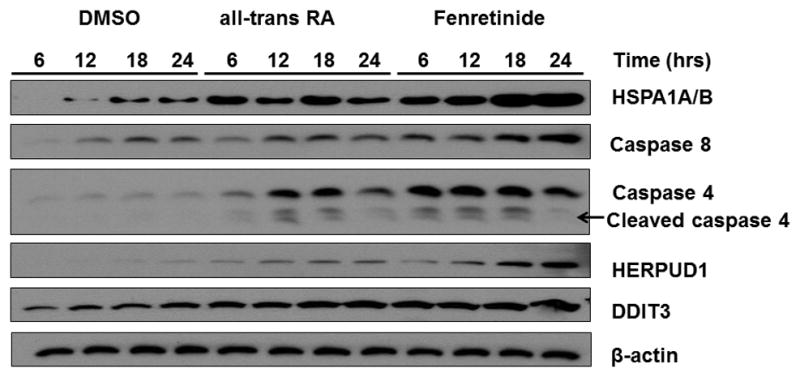
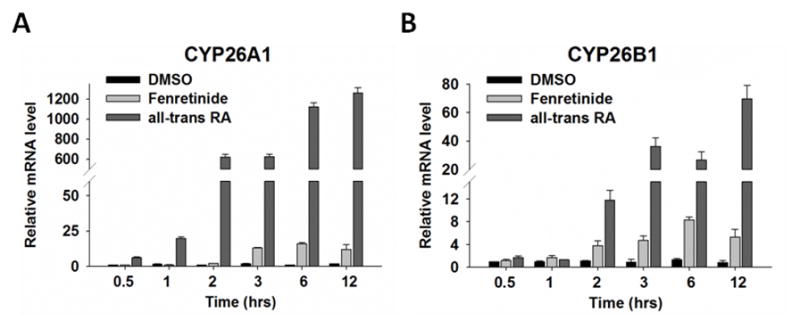
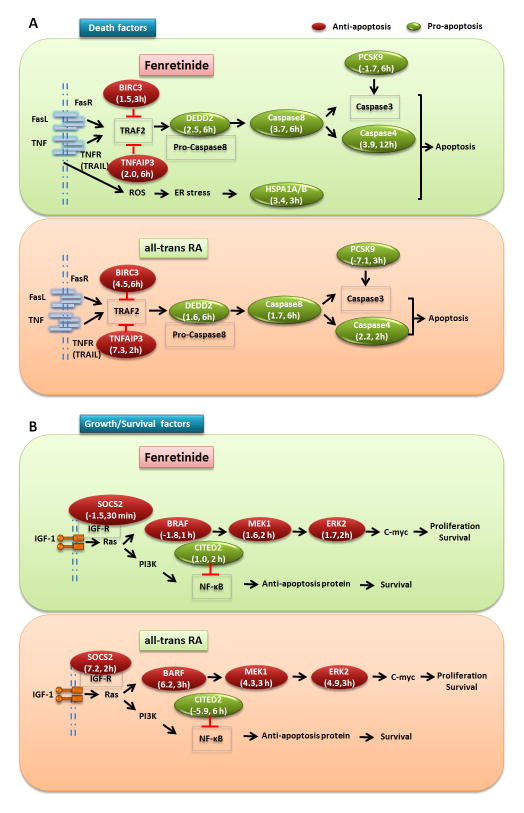
Fenretinide is significantly more effective in inducing apoptosis in cancer cells than all-trans retinoic acid (ATRA). The current study uses a genome-wide approach to understand the differential role fenretinide and ATRA have in inducing apoptosis in Huh7 cells. Fenretinide and ATRA-induced gene expressions and DNA bindings were profiled using microarray and chromatin immunoprecipitation with anti-RXRα antibody. The data showed that fenretinide was not a strong transcription regulator. Fenretinide only changed the expressions of 1 093 genes, approximately three times less than the number of genes regulated by ATRA (2 811). Biological function annotation demonstrated that both fenretinide and ATRA participated in pathways that determine cell fate and metabolic processes. However, fenretinide specifically induced Fas/TNFα-mediated apoptosis by increasing the expression of pro-apoptotic genes i.e., DEDD2, CASP8, CASP4, and HSPA1A/B; whereas, ATRA induced the expression of BIRC3 and TNFAIP3, which inhibit apoptosis by interacting with TRAF2. In addition, fenretinide inhibited the expression of the genes involved in RAS/RAF/ERK-mediated survival pathway. In contrast, ATRA increased the expression of SOSC2, BRAF, MEK, and ERK genes. Most genes regulated by fenretinide and ATRA were bound by RXRα, suggesting a direct effect. This study revealed that by regulating fewer genes, the effects of fenretinide become more specific and thus has fewer side effects than ATRA. The data also suggested that fenretinide induces apoptosis via death receptor effector and by inhibiting the RAS/RAF/ERK pathway. It provides insight on how retinoid efficacy can be improved and how side effects in cancer therapy can be reduced.
Published by Elsevier Inc.
Conflict of interest statement
The authors declare that there are no known conflicts of interest associated with this publication and there has been no significant financial support for this work that could have influenced its outcome.
Figures
Similar articles
-
Biological and clinical implications of retinoic acid-responsive genes in human hepatocellular carcinoma cells.J Hepatol. 2013 Nov;59(5):1037-44. doi: 10.1016/j.jhep.2013.06.024. Epub 2013 Jul 2. J Hepatol. 2013. PMID: 23831118
-
Activation of RAS/ERK alone is insufficient to inhibit RXRα function and deplete retinoic acid in hepatocytes.Biochem Biophys Res Commun. 2014 Sep 26;452(3):801-7. doi: 10.1016/j.bbrc.2014.09.007. Epub 2014 Sep 12. Biochem Biophys Res Commun. 2014. PMID: 25218146
-
Identification of the B-Raf/Mek/Erk MAP kinase pathway as a target for all-trans retinoic acid during skin cancer promotion.Mol Cancer. 2009 May 11;8:27. doi: 10.1186/1476-4598-8-27. Mol Cancer. 2009. PMID: 19432991 Free PMC article.
-
Roles of the Ras/Raf/MEK/ERK pathway in leukemia therapy.Leukemia. 2011 Jul;25(7):1080-94. doi: 10.1038/leu.2011.66. Epub 2011 Apr 15. Leukemia. 2011. PMID: 21494257 Review.
-
Fenretinide and its relation to cancer.Cancer Treat Rev. 1999 Aug;25(4):229-35. doi: 10.1053/ctrv.1999.0127. Cancer Treat Rev. 1999. PMID: 10448131 Review.
Cited by
-
Over-expression of LRIG1 suppresses biological function of pituitary adenoma via attenuation of PI3K/AKT and Ras/Raf/ERK pathways in vivo and in vitro.J Huazhong Univ Sci Technolog Med Sci. 2016 Aug;36(4):558-563. doi: 10.1007/s11596-016-1625-4. Epub 2016 Jul 28. J Huazhong Univ Sci Technolog Med Sci. 2016. PMID: 27465333
-
MiR-22-silenced cyclin A expression in colon and liver cancer cells is regulated by bile acid receptor.J Biol Chem. 2015 Mar 6;290(10):6507-15. doi: 10.1074/jbc.M114.620369. Epub 2015 Jan 17. J Biol Chem. 2015. PMID: 25596928 Free PMC article.
-
Fenretinide: a novel treatment for endometrial cancer.PLoS One. 2014 Oct 23;9(10):e110410. doi: 10.1371/journal.pone.0110410. eCollection 2014. PLoS One. 2014. PMID: 25340777 Free PMC article.
-
Forced expression of fibroblast growth factor 21 reverses the sustained impairment of liver regeneration in hPPARα(PAC) mice due to dysregulated bile acid synthesis.Oncotarget. 2015;6(12):9686-700. doi: 10.18632/oncotarget.3531. Oncotarget. 2015. PMID: 25991671 Free PMC article.
-
miR-22 gene therapy treats HCC by promoting anti-tumor immunity and enhancing metabolism.Mol Ther. 2023 Jun 7;31(6):1829-1845. doi: 10.1016/j.ymthe.2023.04.019. Epub 2023 May 4. Mol Ther. 2023. PMID: 37143325 Free PMC article.
References
-
- Choi Y, Kim SY, Kim SH, Yang J, Park K, Byun Y. Inhibition of tumor growth by biodegradable microspheres containing all-trans-retinoic acid in a human head-and-neck cancer xenograft. Int J Cancer. 2003;107:145–8. - PubMed
-
- Di C, Liao S, Adamson DC, Parrett TJ, Broderick DK, Shi Q, et al. Identification of OTX2 as a medulloblastoma oncogene whose product can be targeted by all-trans retinoic acid. Cancer Res. 2005;65:919–24. - PubMed
-
- de-Medeiros BC, Strapasson E, Pasquini R, de-Medeiros CR. Effect of all-trans retinoic acid on newly diagnosed acute promyelocytic leukemia patients: results of a Brazilian center. Braz J Med Biol Res. 1998;31:1537–43. - PubMed
-
- Tari AM, Lim SJ, Hung MC, Esteva FJ, Lopez-Berestein G. Her2/neu induces all-trans retinoic acid (ATRA) resistance in breast cancer cells. Oncogene. 2002;21:5224–32. - PubMed
-
- Kitareewan S, Spinella MJ, Allopenna J, Reczek PR, Dmitrovsky E. 4HPR triggers apoptosis but not differentiation in retinoid sensitive and resistant human embryonal carcinoma cells through an RARγ independent pathway. Oncogene. 1999;18:5747–55. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
