

Endogenous retroviruses function as species-specific enhancer elements in the placenta
- PMID: 23396136
- PMCID: PMC3789077
- DOI: 10.1038/ng.2553
Endogenous retroviruses function as species-specific enhancer elements in the placenta
Abstract
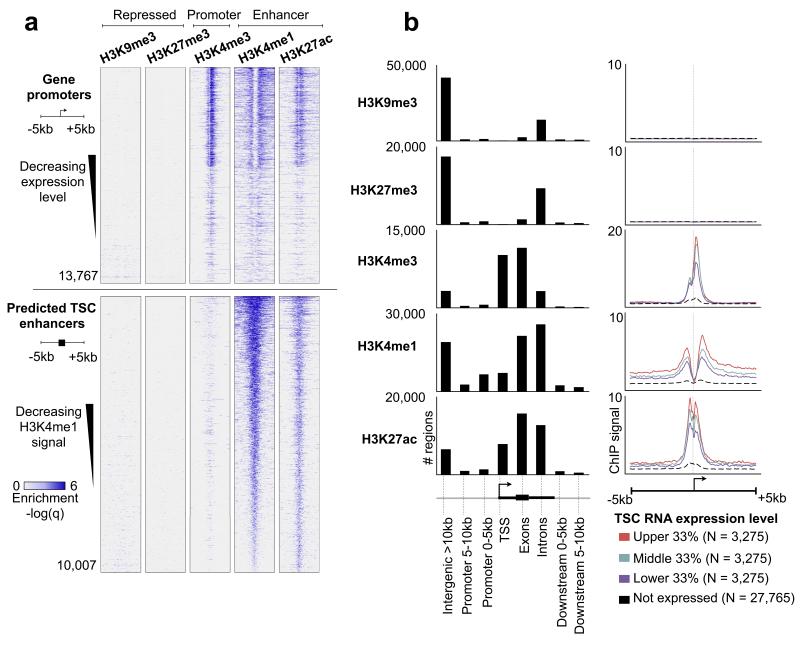
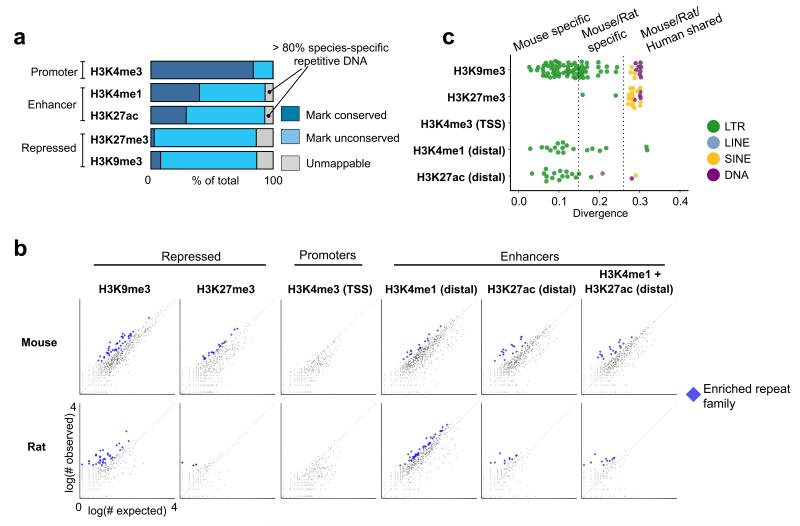
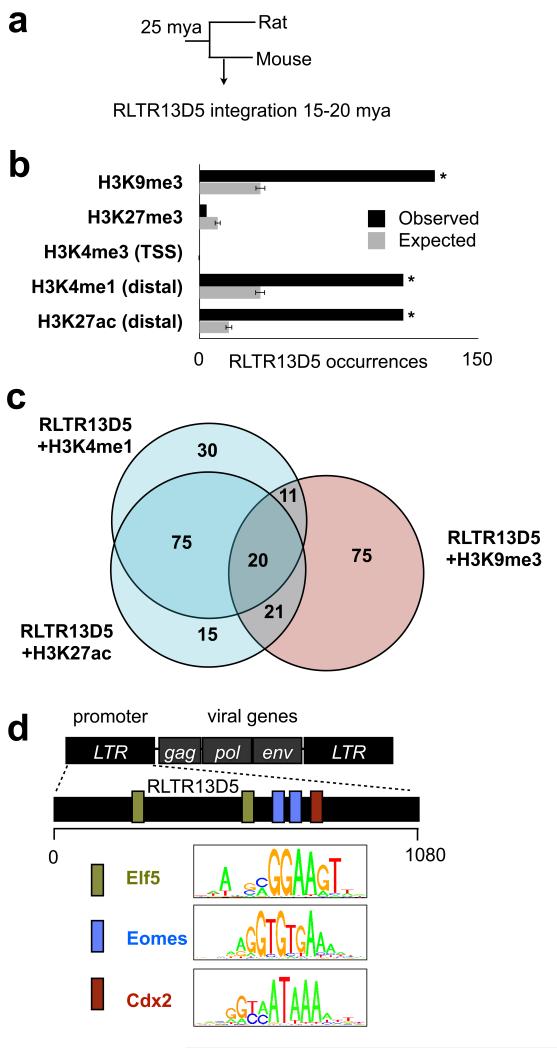
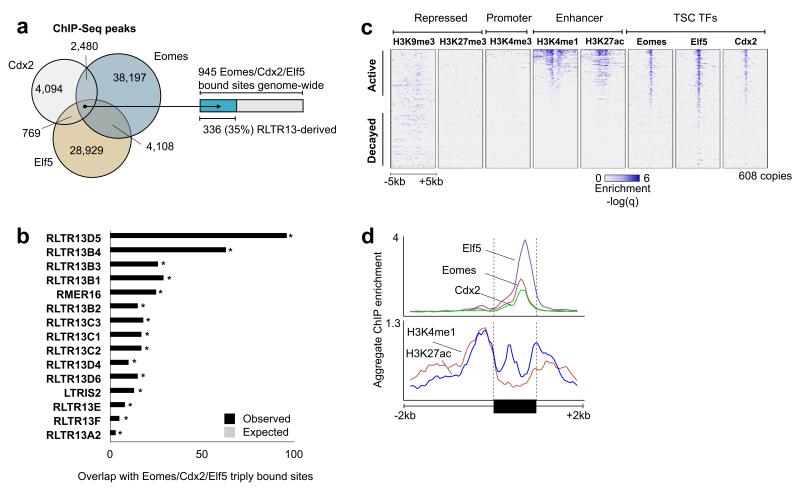
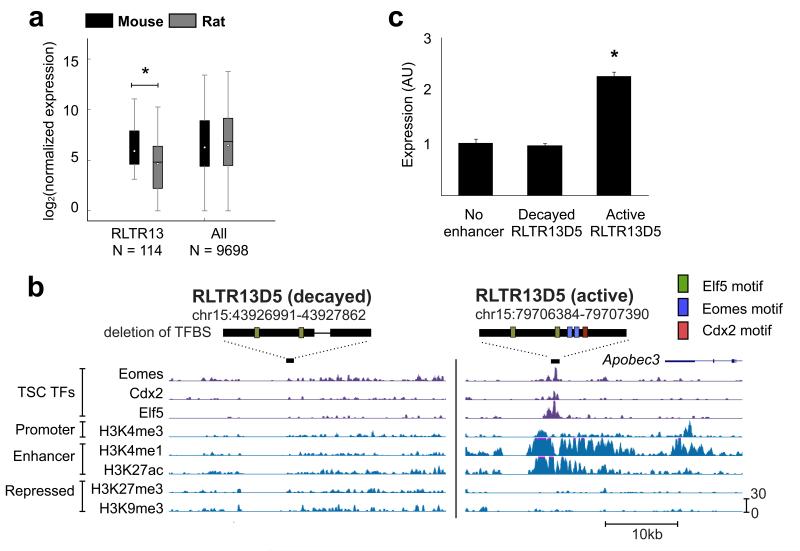
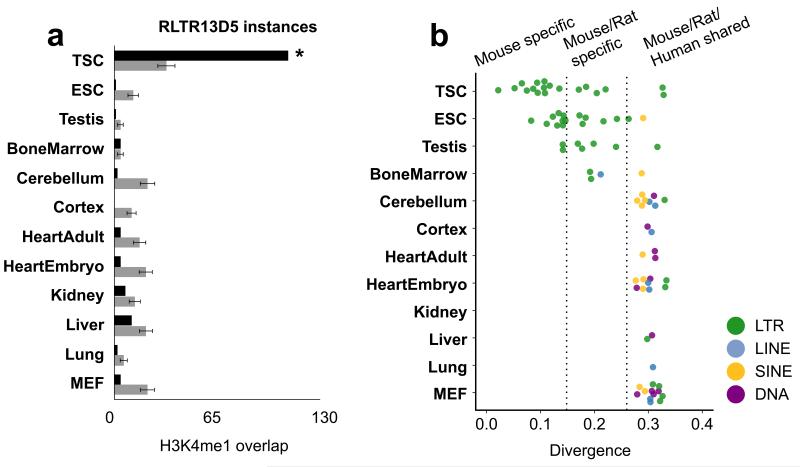
The mammalian placenta is remarkably distinct between species, suggesting a history of rapid evolutionary diversification. To gain insight into the molecular drivers of placental evolution, we compared biochemically predicted enhancers in mouse and rat trophoblast stem cells (TSCs) and found that species-specific enhancers are highly enriched for endogenous retroviruses (ERVs) on a genome-wide level. One of these ERV families, RLTR13D5, contributes hundreds of mouse-specific histone H3 lysine 4 monomethylation (H3K4me1)- and histone H3 lysine 27 acetylation (H3K27ac)-defined enhancers that functionally bind Cdx2, Eomes and Elf5-core factors that define the TSC regulatory network. Furthermore, we show that RLTR13D5 is capable of driving gene expression in rat placental cells. Analysis in other tissues shows that species-specific ERV enhancer activity is generally restricted to hypomethylated tissues, suggesting that tissues permissive for ERV activity gain access to an otherwise silenced source of regulatory variation. Overall, our results implicate ERV enhancer co-option as a mechanism underlying the extensive evolutionary diversification of placental development.
Figures
References
-
- Mossman HW. Vertebrate Fetal Membranes: Comparative Ontogeny and Morphology; Evolution; Phylogenetic Significance; Basic Functions; Research Opportunities. Rutgers University Press; 1987.
-
- Enders A. Reasons for Diversity of Placental Structure. Placenta. 2009;30(Supplement):15–18. - PubMed
-
- Crespi B, Semeniuk C. Parent-offspring conflict in the evolution of vertebrate reproductive mode. Am Nat. 2004;163:635–653. - PubMed
-
- Haig D. Genetic conflicts in human pregnancy. The Quarterly Review of Biology. 1993;68:495–532. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
