

Ketone body metabolism and cardiovascular disease
- PMID: 23396451
- PMCID: PMC3625904
- DOI: 10.1152/ajpheart.00646.2012
Ketone body metabolism and cardiovascular disease
Abstract
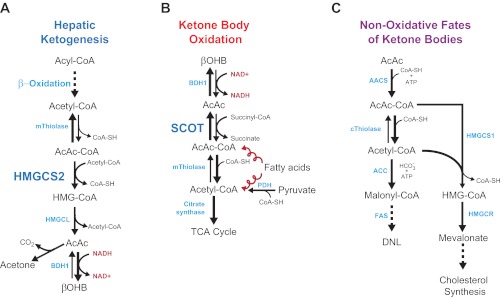
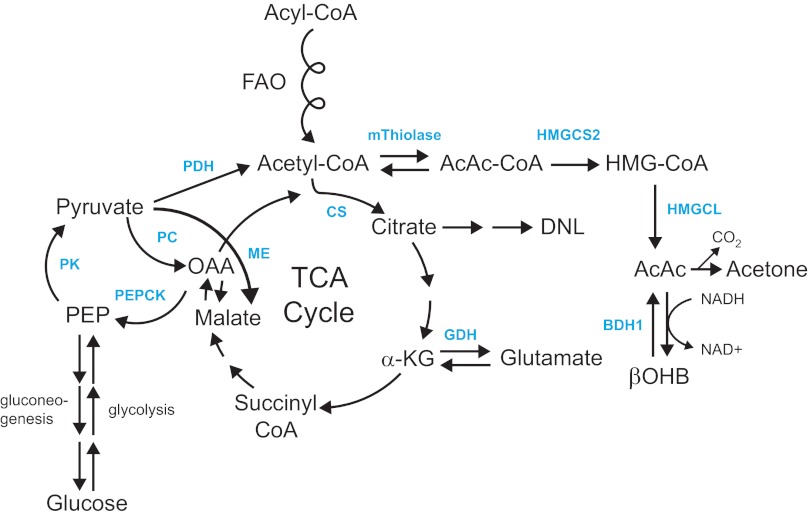
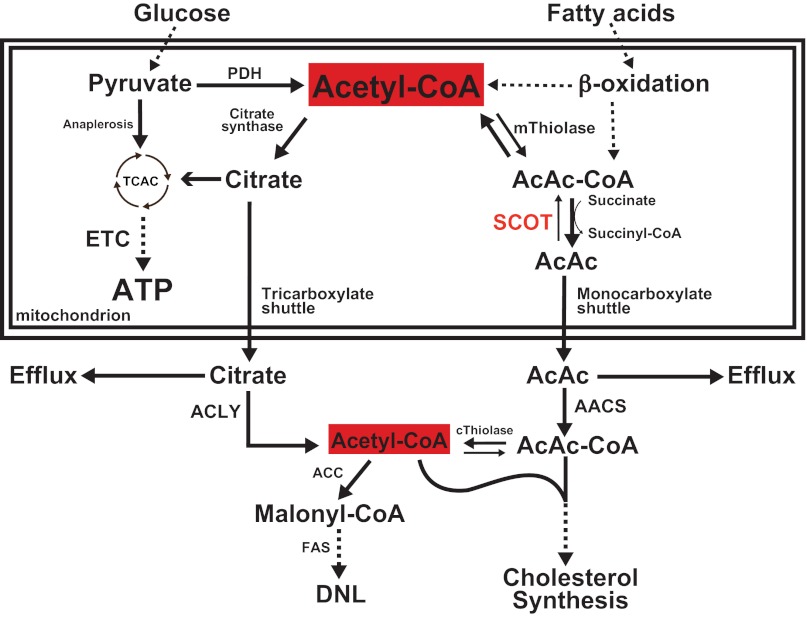
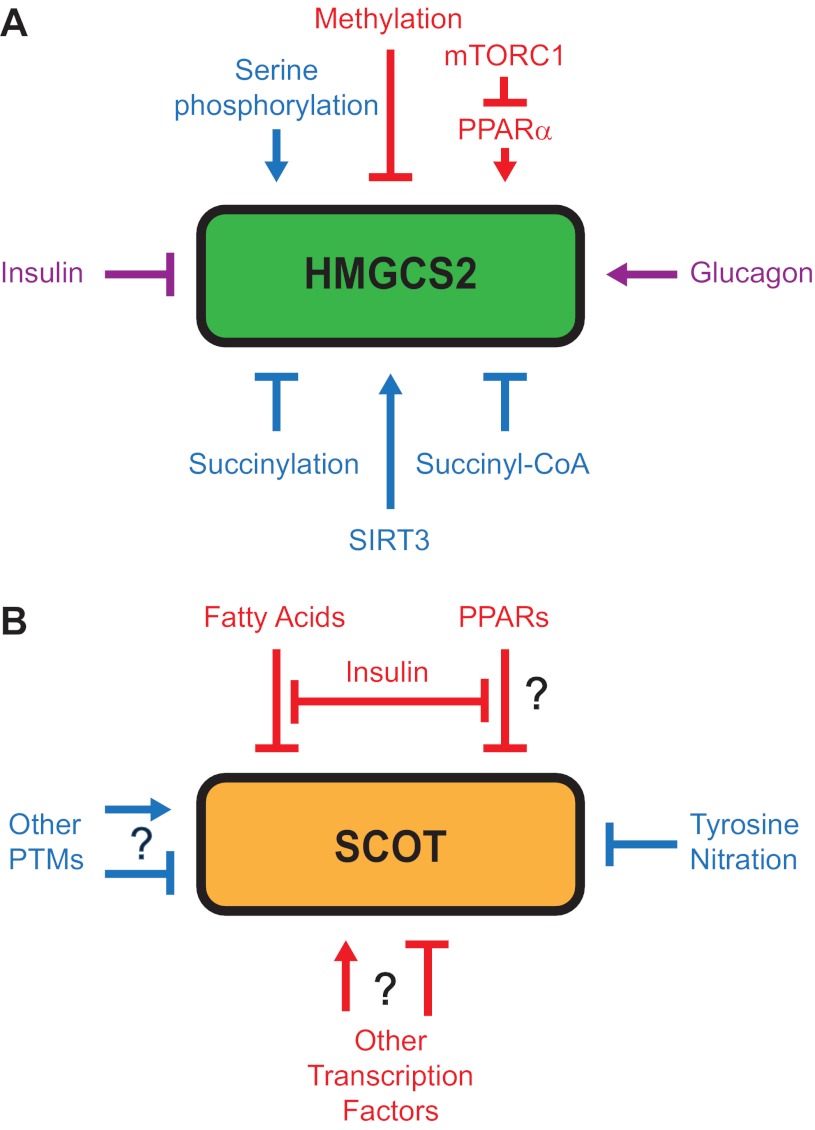
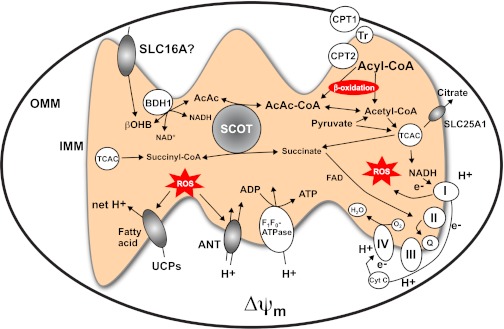
Ketone bodies are metabolized through evolutionarily conserved pathways that support bioenergetic homeostasis, particularly in brain, heart, and skeletal muscle when carbohydrates are in short supply. The metabolism of ketone bodies interfaces with the tricarboxylic acid cycle, β-oxidation of fatty acids, de novo lipogenesis, sterol biosynthesis, glucose metabolism, the mitochondrial electron transport chain, hormonal signaling, intracellular signal transduction pathways, and the microbiome. Here we review the mechanisms through which ketone bodies are metabolized and how their signals are transmitted. We focus on the roles this metabolic pathway may play in cardiovascular disease states, the bioenergetic benefits of myocardial ketone body oxidation, and prospective interactions among ketone body metabolism, obesity, metabolic syndrome, and atherosclerosis. Ketone body metabolism is noninvasively quantifiable in humans and is responsive to nutritional interventions. Therefore, further investigation of this pathway in disease models and in humans may ultimately yield tailored diagnostic strategies and therapies for specific pathological states.
Figures
References
-
- Aguilo F, Camarero N, Relat J, Marrero PF, Haro D. Transcriptional regulation of the human acetoacetyl-CoA synthetase gene by PPARgamma. Biochem J 427: 255–264, 2010 - PubMed
-
- Ahmed K, Tunaru S, Offermanns S. GPR109A, GPR109B and GPR81, a family of hydroxy-carboxylic acid receptors. Trends Pharmacol Sci 30: 557–562, 2009 - PubMed
-
- Al-Zaid NS, Dashti HM, Mathew TC, Juggi JS. Low carbohydrate ketogenic diet enhances cardiac tolerance to global ischaemia. Acta Cardiol 62: 381–389, 2007 - PubMed
-
- Aledo R, Zschocke J, Pie J, Mir C, Fiesel S, Mayatepek E, Hoffmann GF, Casals N, Hegardt FG. Genetic basis of mitochondrial HMG-CoA synthase deficiency. Hum Genet 109: 19–23, 2001 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases