

Identification and characterization of MalA in the maltose/maltodextrin operon of Sulfolobus acidocaldarius DSM639
- PMID: 23396915
- PMCID: PMC3624569
- DOI: 10.1128/JB.01713-12
Identification and characterization of MalA in the maltose/maltodextrin operon of Sulfolobus acidocaldarius DSM639
Abstract
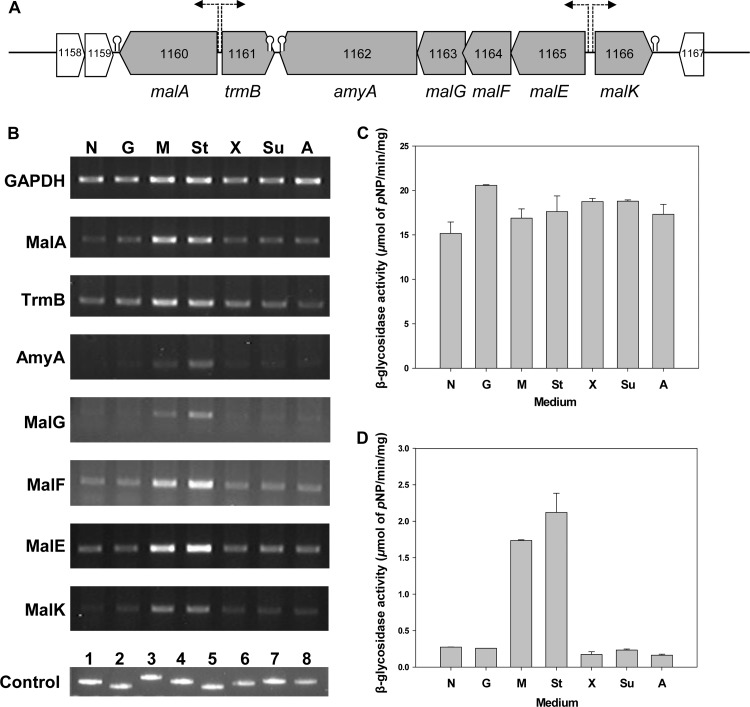
A putative maltose/maltodextrin operon was found in the Sulfolobus acidocaldarius DSM639 genome. The gene cluster consisted of 7 genes (malA, trmB, amyA, malG, malF, malE, and malK). Here, we report the identification of MalA, which is responsible for the hydrolysis of maltose or maltodextrin to glucose in S. acidocaldarius. The transcription level of malA was increased 3-fold upon the addition of maltose or starch to the medium. Moreover, the α-glucosidase activity for maltose as a substrate in cell extracts of S. acidocaldarius DSM639 was also 11- and 10-fold higher during growth in YT medium (Brock's mineral salts, 0.1% [wt/vol] tryptone, and 0.005% [wt/vol] yeast extract) containing maltose or starch, respectively, than during growth on other sugars. The gene encoding MalA was cloned and expressed in S. acidocaldarius. The enzyme purified from the organism was a dodecamer in its active state and showed strong maltose-hydrolyzing activity at 100°C and pH 5.0. MalA was remarkably thermostable, with half-lives of 33.8 h, 10.6 h, and 1.8 h at 95°C, 100°C, and 105°C, respectively. Substrate specificity and kinetic studies of MalA with maltooligosaccharides indicated that MalA efficiently hydrolyzed maltose to maltopentaose, which is a typical characteristic of GH31-type α-glucosidases. However, glycogen or starch was not hydrolyzed. Reverse transcription-PCR, sugar uptake, and growth studies of the wild-type DSM639 and ΔmalEFG mutant on different sugars demonstrated that MalA located in the mal operon gene cluster is involved in maltose and starch metabolism in S. acidocaldarius.
Figures






References
-
- Berkner S, Lipps G. 2008. Genetic tools for Sulfolobus spp.: vectors and first applications. Arch. Microbiol. 190:217–230 - PubMed
-
- Elferink MG, Albers SV, Konings WN, Driessen AJM. 2001. Sugar transport in Sulfolobus solfataricus is mediated by two families of binding protein-dependent ABC transporters. Mol. Microbiol. 39:1494–1503 - PubMed
-
- Lubelska JM, Jonuscheit M, Schleper C, Albers SV, Driessen AJ. 2006. Regulation of expression of the arabinose and glucose transporter genes in the thermophilic archaeon Sulfolobus solfataricus. Extremophiles 10:383–391 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
