

Review
doi: 10.1038/nrg3375.
Epub 2013 Feb 12.
Organization and segregation of bacterial chromosomes
Affiliations
- PMID: 23400100
- PMCID: PMC3869393
- DOI: 10.1038/nrg3375
Item in Clipboard
Review
Organization and segregation of bacterial chromosomes
Nat Rev Genet.
2013 Mar.
Abstract
The bacterial chromosome must be compacted more than 1,000-fold to fit into the compartment in which it resides. How it is condensed, organized and ultimately segregated has been a puzzle for over half a century. Recent advances in live-cell imaging and genome-scale analyses have led to new insights into these problems. We argue that the key feature of compaction is the orderly folding of DNA along adjacent segments and that this organization provides easy and efficient access for protein-DNA transactions and has a central role in driving segregation. Similar principles and common proteins are used in eukaryotes to condense and to resolve sister chromatids at metaphase.
Figures
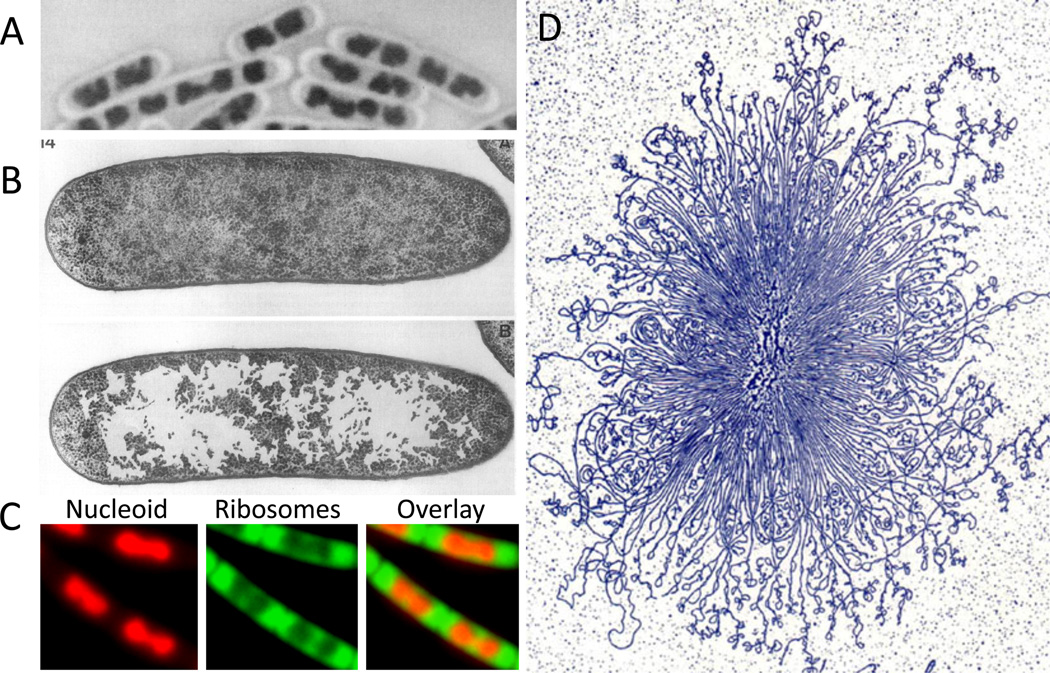
(A) B. subtilis nucleoid stained with Giemsa using acid-treated cells. (B) The nucleoid of growing E. coli in thin section after cryo-fixation followed by freeze-substitution. The upper and lower panels show the same section; in the lower panel, the ribosome-free spaces were enhanced by coloring by hand. (A) and (B) are adapted from Robinow and Kellenberger . (C) Nucleoid (stained with DAPI, colored red) and ribosomes (RplA-GFP, colored green) in live B. subtilis cells growing in rich media. Despite this commonly depicted cloud-like appearance of the bacterial chromosome, the morphology of the nucleoid varies among bacteria, and is influenced by growth rate and environmental conditions. For example, the nucleoid in C. crescentus, and in slow-growing E. coli and B. subtilis, appears more diffuse and occupies a greater proportion of the cell cytoplasm (not shown). (D) A gently isolated E. coli nucleoid bound by cytochrome C, spread on an EM grid, stained with uranyl acetate and visualized by transmission electron microscopy. Adapted from Physics in the twentieth century .
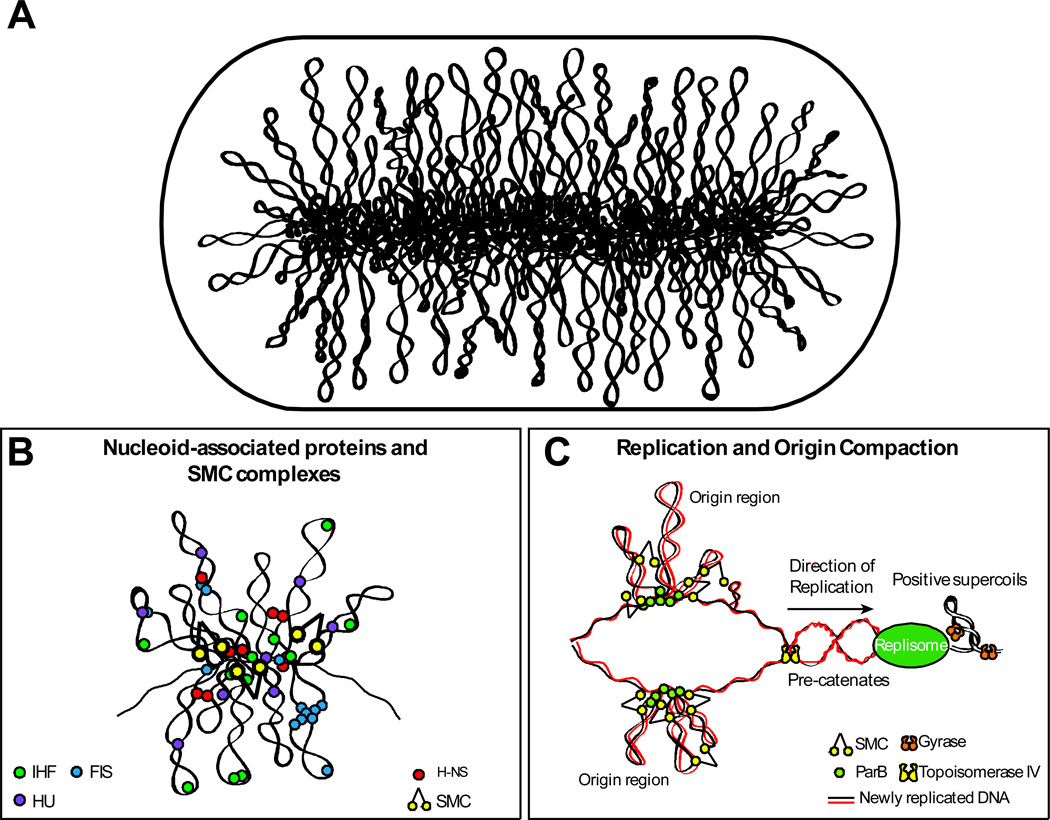
(A) Schematic representation of the bottlebrush model of the nucleoid. This diagram depicts the interwound supercoiled loops emanating from a dense core. The topologically isolated domains (microdomains) are on average 10 kb and therefore likely encompass several branched plectonemic loops. (B) Schematic representation of the small nucleoid-associated proteins and SMC. These proteins introduce DNA bends and also function in bridging chromosomal loci. (C) The diagram depicts replication fork progression and compaction of the origin region. Replication generates positive supercoils ahead of the fork, which can diffuse behind the replisome producing pre-catenanes. Positive supercoils are removed by DNA gyrase and pre-catenanes are unlinked by Topo IV. Newly replicated origin regions thought to be compacted by the SMC complexes that are recruited to the origin and by the action of small nucleoid-associated proteins (not shown).
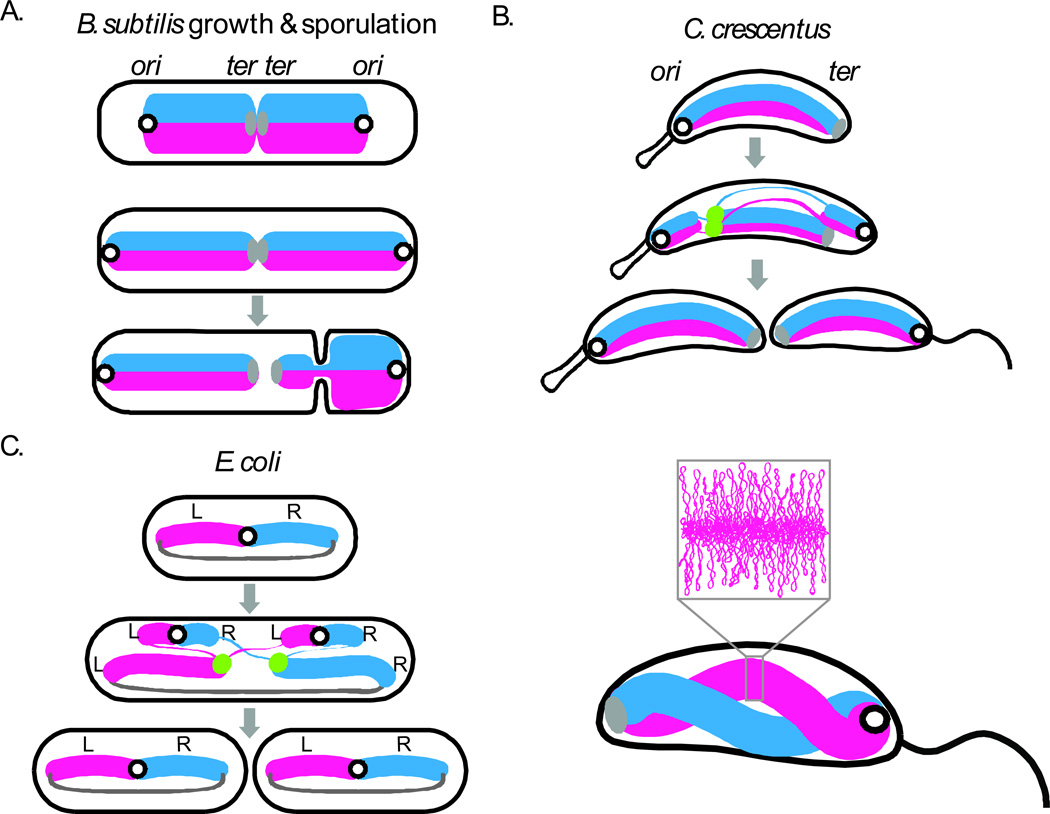
(A) Cellular organization of the chromosome in B. subtilis during growth and spore formation, C. crescentus (B) and slow growing E. coli (C) during vegetative cell cycles (gray arrows). During sporulation in B. subtilis, an asymmetric division traps about 25% of the chromosome in the smaller spore compartment ,. A DNA translocase (not shown) pumps the remaining 75% of the chromosome into spore after cytokinesis . The lower panel of (B) shows a model for the spatially separate but gently twisted arms of the C. crescentus chromosome based on chromosome conformation capture (new ref). The inset shows the plectonemic loops within one of the arms. Replication origins are illustrated as black open circles, termini as solid gray ovals (or lines in E. coli), the replisomes as green dots, the left (L) and right (R) replication arms as thick blue and pink lines, and newly replicated DNA as thinner lines. This figure is adapted from Jackson et al (2012).
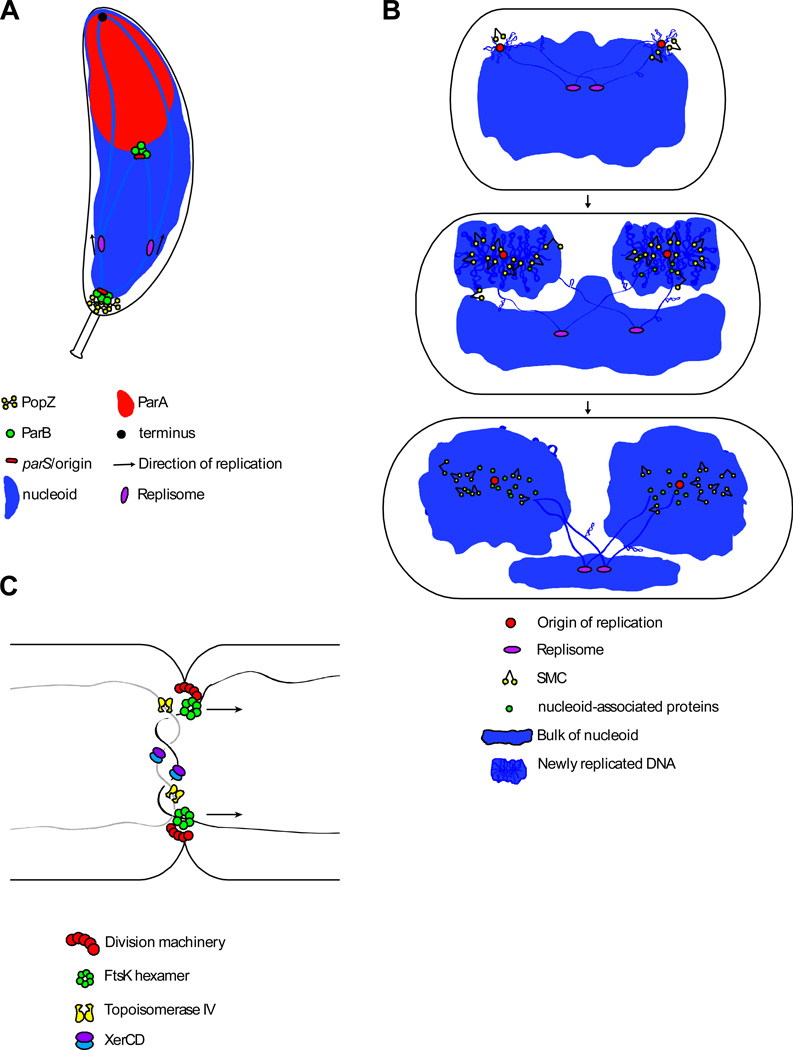
(A) Schematic model of Par-mediated origin segregation in C. crescentus–. The origin region is tethered to the cell pole through interactions of ParB/parS with the polar anchor PopZ. After initiation of replication, one of the sister origins is pulled toward the opposite pole through interactions between ParB/parS and ParA(ATP) bound non-specifically to the nucleoid. These interactions trigger hydrolysis of ParA(ATP) and release of ParA(ADP) from the nucleoid. The ParB/pars complex then binds to neighboring ParA(ATP) on the nucleoid (alternatively, another ParB in the nucleoprotein complex engaged a nearby ParA(ATP) prior to release of the first). Repeated cycles of binding, hydrolysis, and release results in movement of the ParB/pars complex toward the cell pole and a ParA-free nucleoid in its wake. This so-called diffusion-ratchet mechanism allows the ParB/parS complex to “surf” on top of the nucleoid toward the pole ,. In C. crescentus, an additional protein (TipN) located at the cell pole is required for Par-mediated segregation , (not shown). TipN localizes to the new cell pole where it likely functions to regenerate ParA(ATP) helping to set up a ParA(ATP) gradient on the nucleoid. In par-containing bacteria that do not anchor their origins at the cell pole, the partitioning system helps re-position the newly replicated origins at the polar edges of the nucleoid. (B) Schematic model of bulk chromosome segregation. After newly replicated origins are separated, lengthwise condensation mediated by supercoiling, small nucleoid-associated proteins and SMC, in lock-step with replication, drive disentanglement and segregation of the sister chromosomes. (C) Schematic of terminus segregation in E. coli. The replicated terminus is translocated to appropriate daughter cell by the FtsK DNA translocase, while Topoisomerase IV and XerCD resolve catenanes and chromosome dimers, respectively. FtsK specifically localizes at the division septum where it participates in cytokinesis and DNA segregation.
References
-
- Baker JR. The cell-theory: a restatement, history and critique. Part V. The multiplication of nuclei. Q J Microsc Sci. 1955;96:449–481.
-
- Piekarski G. Zytologische Untersuchungen an Paratyphusund Coli Bakterien. Arch. Mikrobiol. 1937;8:428–429.
-
- Stille B. Zytologische Untersuchungen an Bakterien mit Hilfe der Feulgenschen Nuclealreaktion. Arch. Mikrobiol. 1937;8:124–148.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
