

Deficiency for endoglin in tumor vasculature weakens the endothelial barrier to metastatic dissemination
- PMID: 23401487
- PMCID: PMC3600899
- DOI: 10.1084/jem.20120662
Deficiency for endoglin in tumor vasculature weakens the endothelial barrier to metastatic dissemination
Abstract
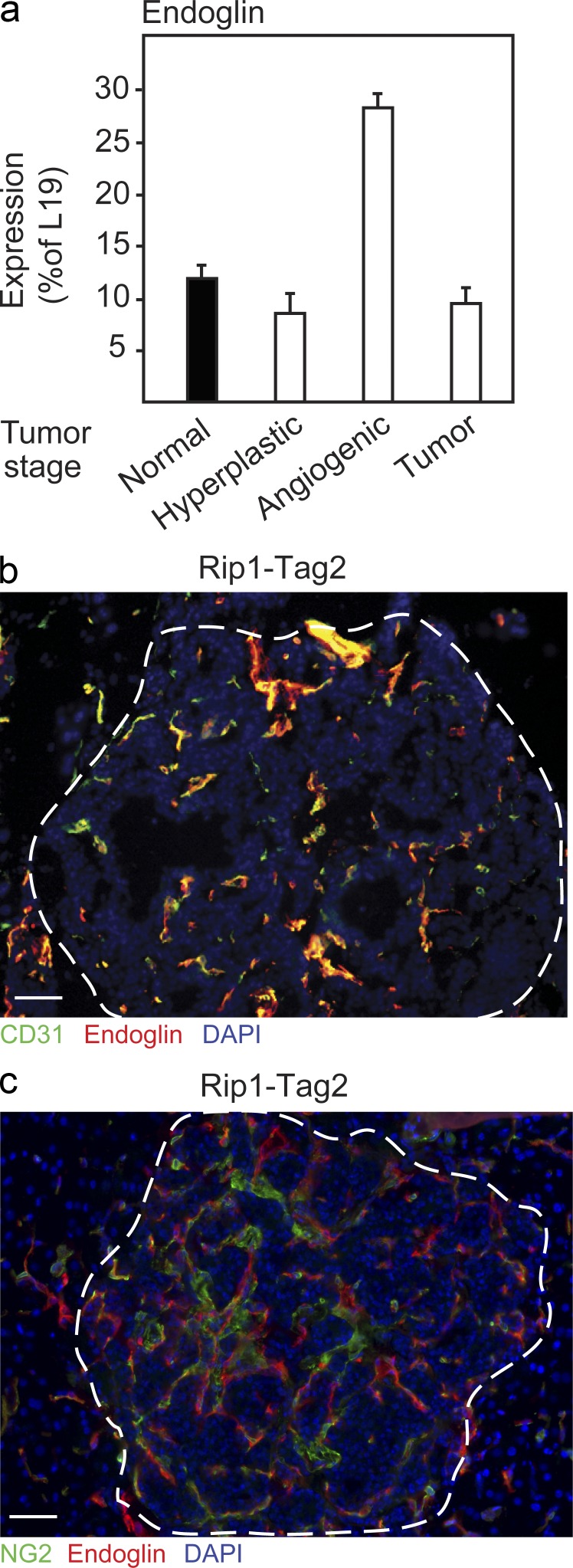
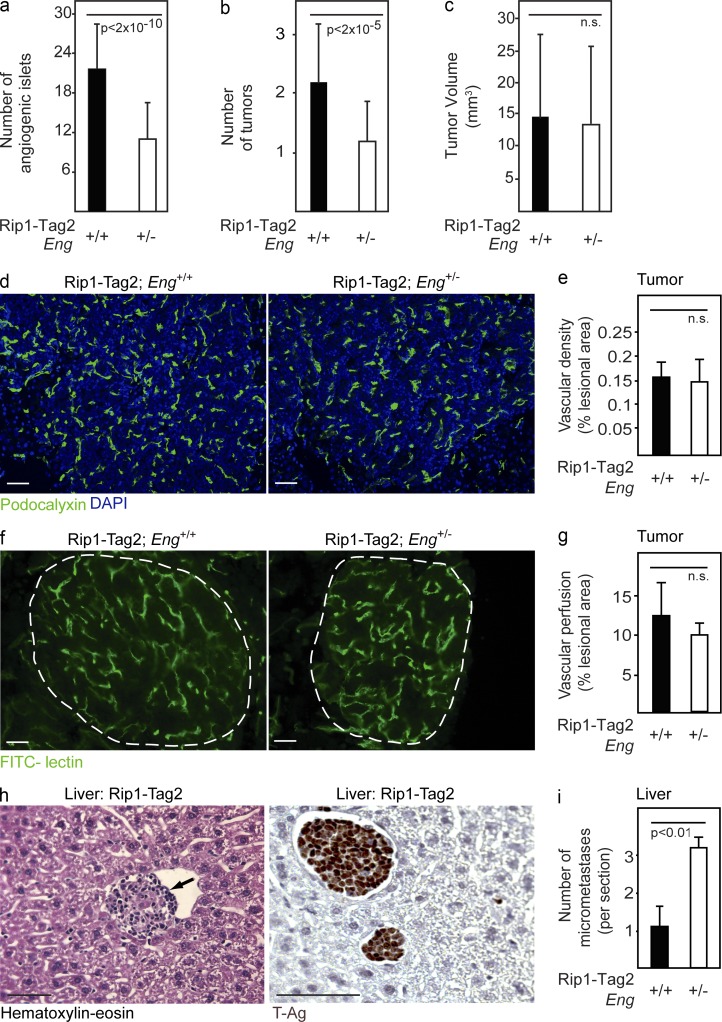
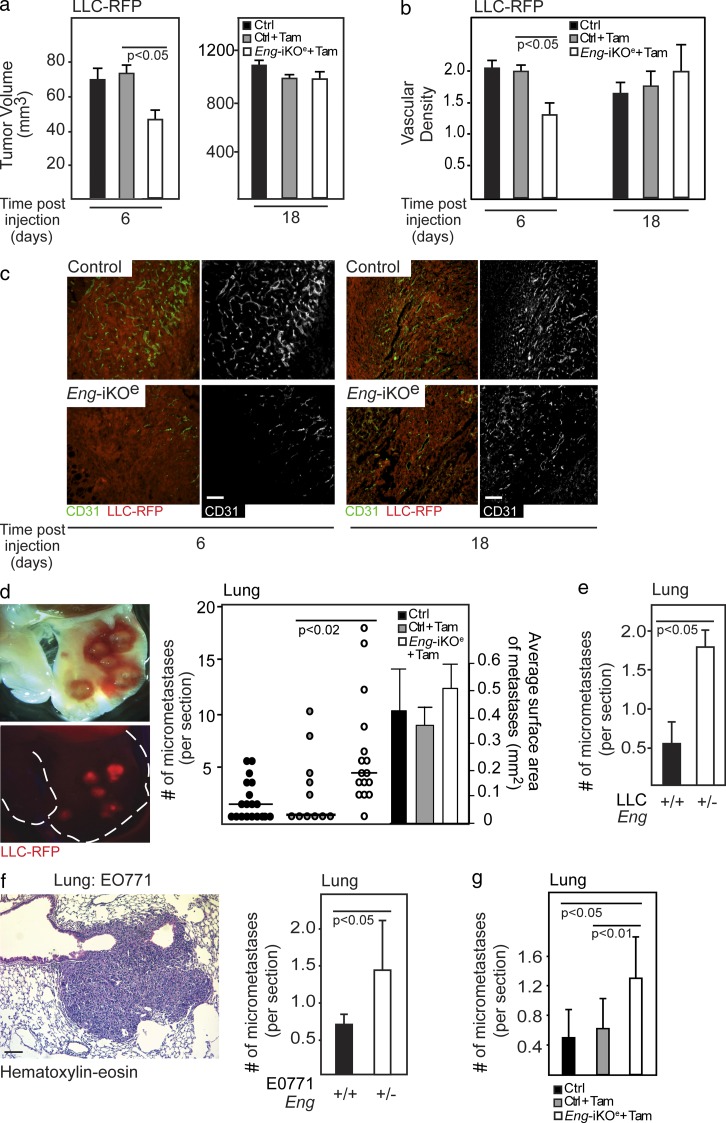
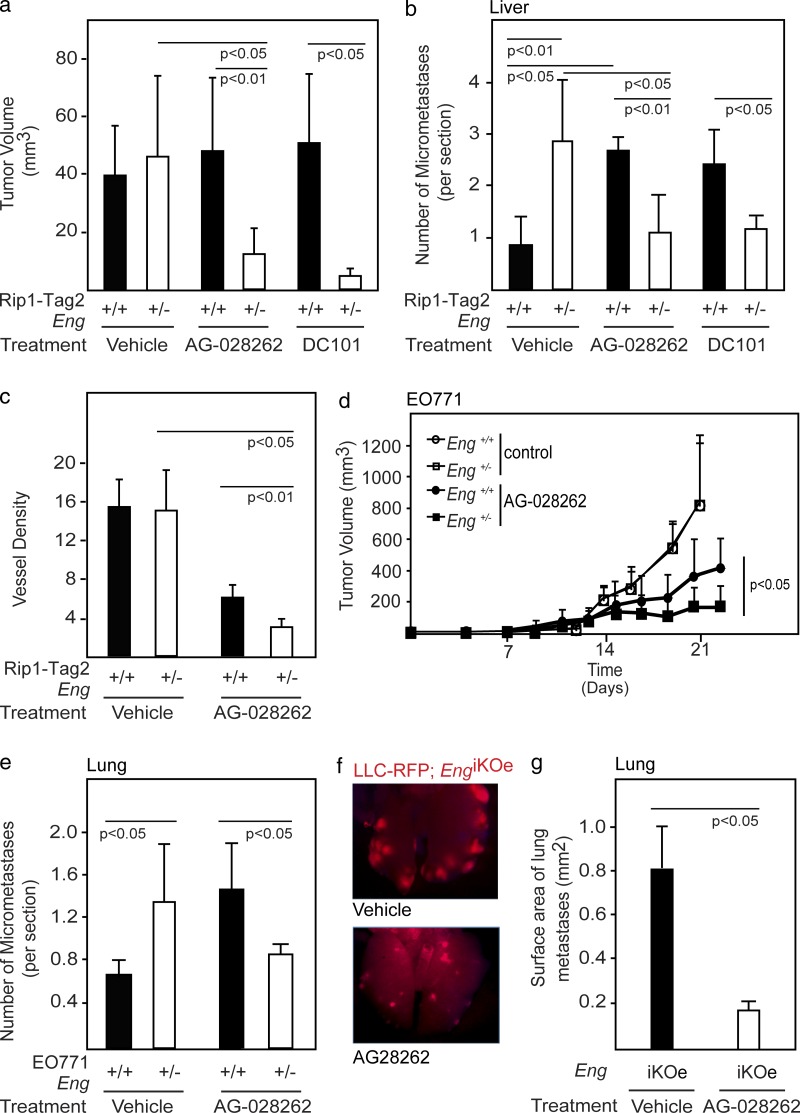
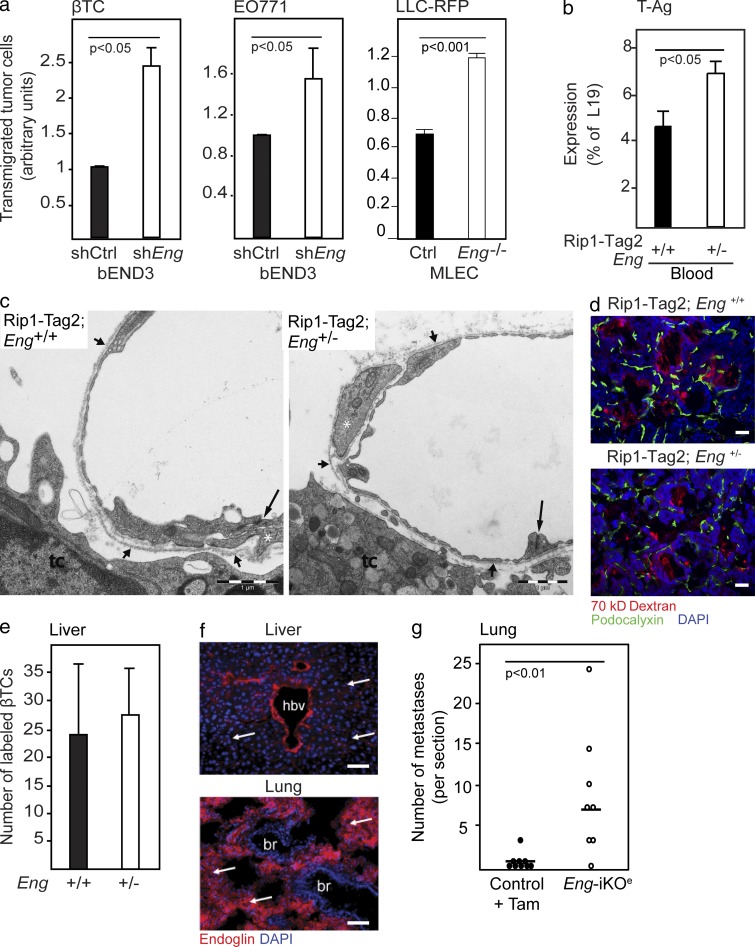
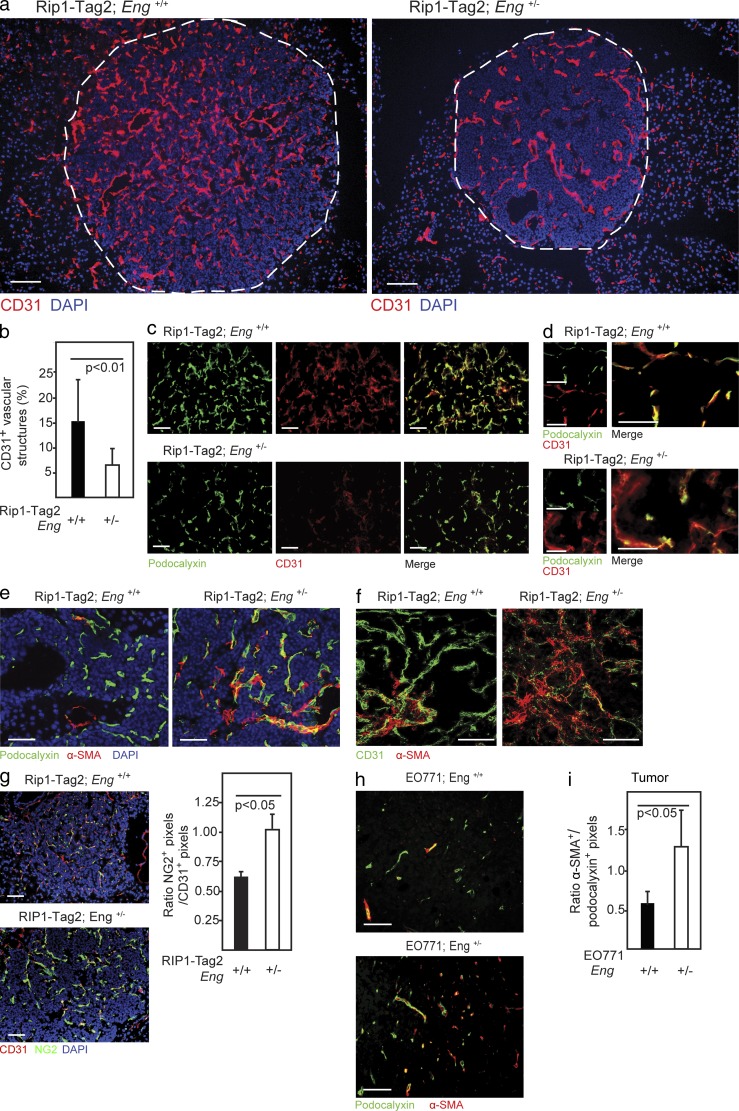
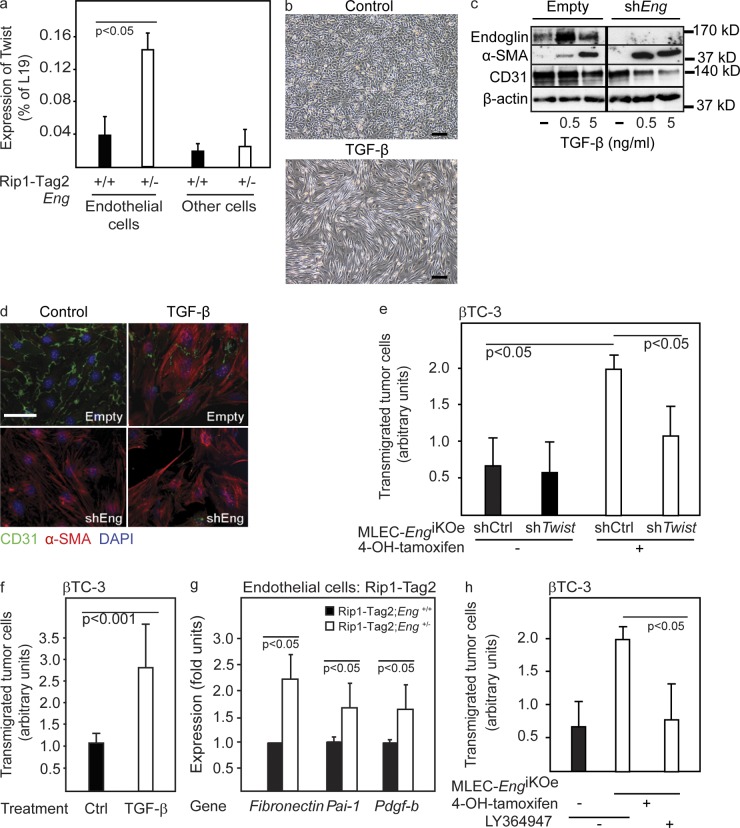
Therapy-induced resistance remains a significant hurdle to achieve long-lasting responses and cures in cancer patients. We investigated the long-term consequences of genetically impaired angiogenesis by engineering multiple tumor models deprived of endoglin, a co-receptor for TGF-β in endothelial cells actively engaged in angiogenesis. Tumors from endoglin-deficient mice adapted to the weakened angiogenic response, and refractoriness to diminished endoglin signaling was accompanied by increased metastatic capability. Mechanistic studies in multiple mouse models of cancer revealed that deficiency for endoglin resulted in a tumor vasculature that displayed hallmarks of endothelial-to-mesenchymal transition, a process of previously unknown significance in cancer biology, but shown by us to be associated with a reduced capacity of the vasculature to avert tumor cell intra- and extravasation. Nevertheless, tumors deprived of endoglin exhibited a delayed onset of resistance to anti-VEGF (vascular endothelial growth factor) agents, illustrating the therapeutic utility of combinatorial targeting of multiple angiogenic pathways for the treatment of cancer.
Figures
References
-
- Arthur H.M., Ure J., Smith A.J., Renforth G., Wilson D.I., Torsney E., Charlton R., Parums D.V., Jowett T., Marchuk D.A., et al. 2000. Endoglin, an ancillary TGFbeta receptor, is required for extraembryonic angiogenesis and plays a key role in heart development. Dev. Biol. 217:42–53 10.1006/dbio.1999.9534 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
