

Perilipin-2-null mice are protected against diet-induced obesity, adipose inflammation, and fatty liver disease
- PMID: 23402988
- PMCID: PMC3622329
- DOI: 10.1194/jlr.M035063
Perilipin-2-null mice are protected against diet-induced obesity, adipose inflammation, and fatty liver disease
Abstract
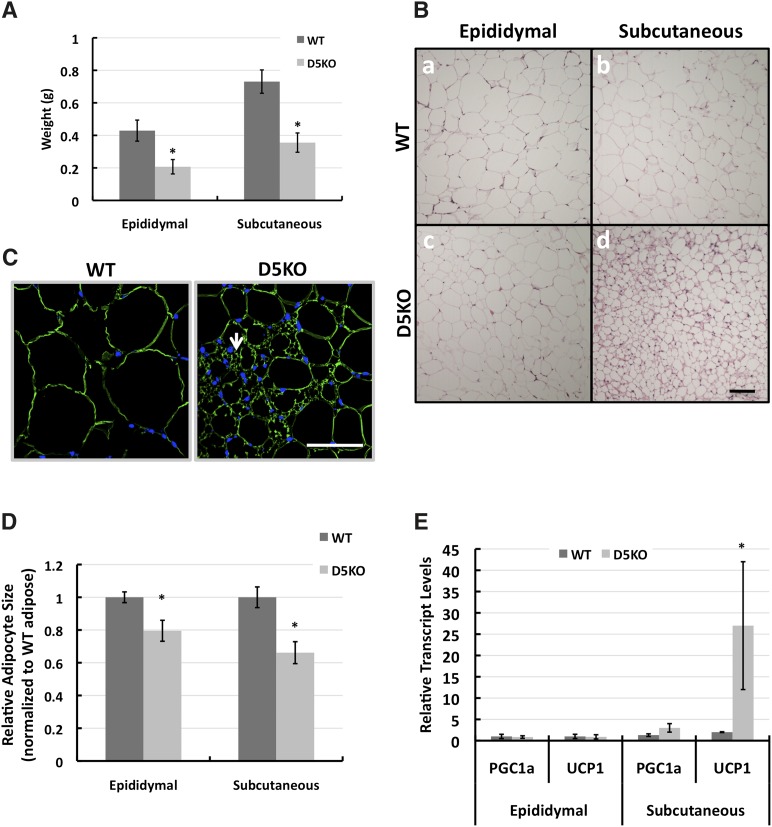
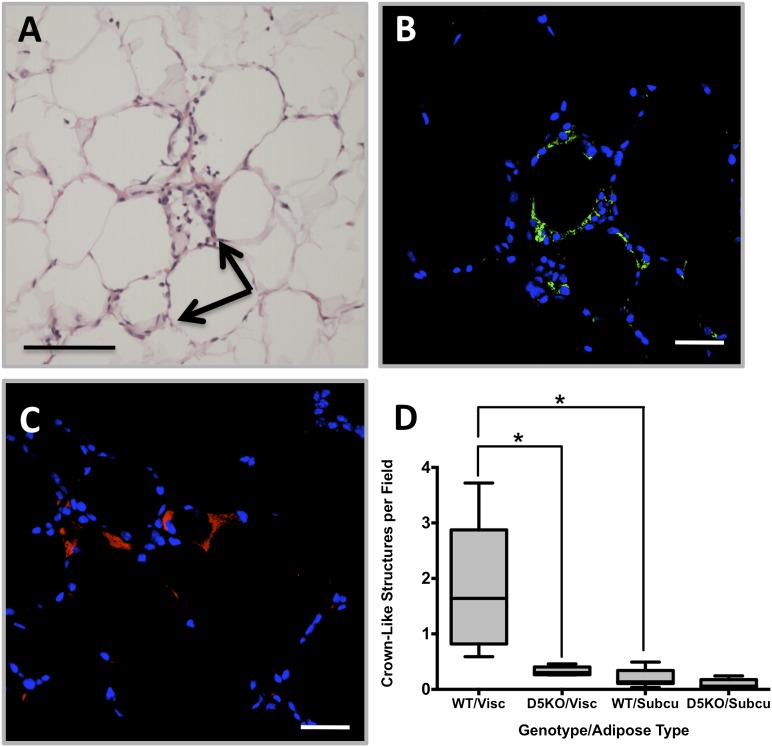
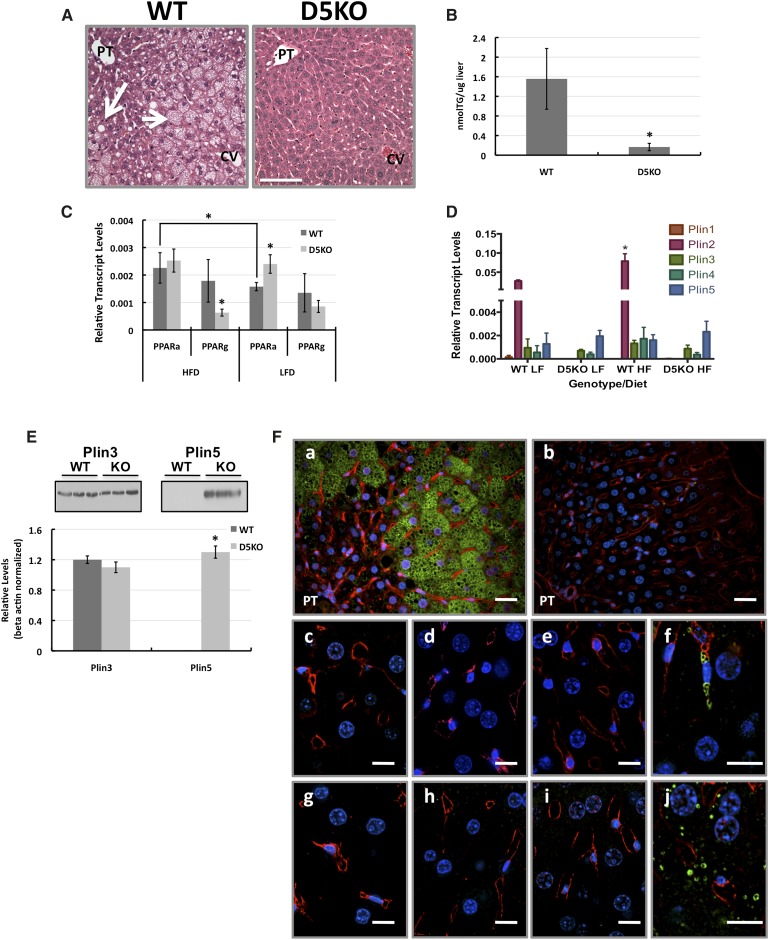
The cytoplasmic lipid droplet (CLD) protein perilipin-2 (Plin2) is expressed in multiple nonadipose tissues, where it is thought to play a role in regulating their lipid storage properties. However, the extent to which Plin2 functions in nutrient utilization and metabolism, or how it influences the consequences of over-feeding, remains unclear. In this study, we demonstrate that the absence of Plin2 prevents high-fat diet(HFD)-induced obesity in male and female mice. This response is associated with increased formation of subcutaneous beige adipocyte cells with uncoupling protein 1 expression, and amelioration of inflammatory foci formation in white adipose tissue and steatosis in the liver. Experiments demonstrate that Plin2 loss results in reduced energy intake and increased physical activity in response to HFD feeding. Our study provides the first evidence that Plin2 contributes to HFD-induced obesity by modulating food intake, and that its absence prevents obesity-associated adipose tissue inflammatory foci and liver steatosis.
Figures
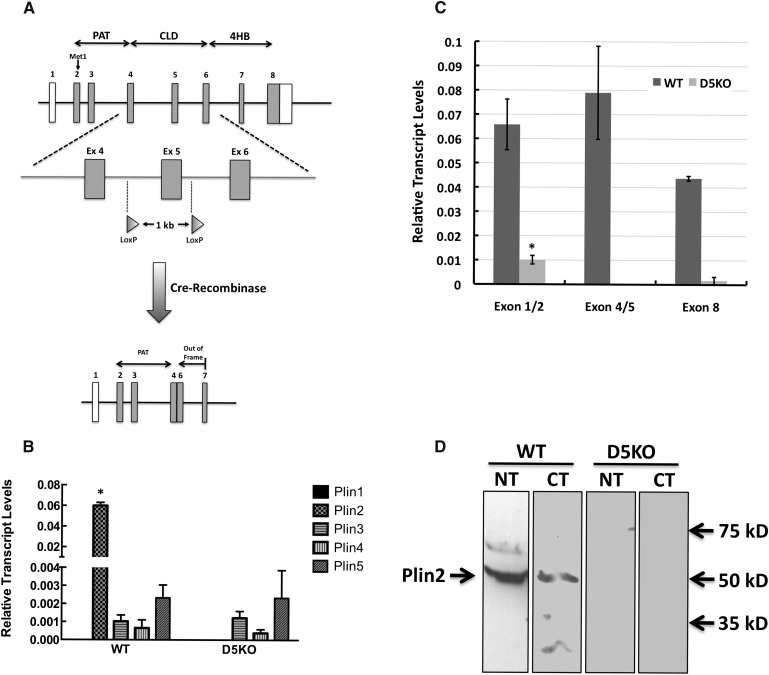
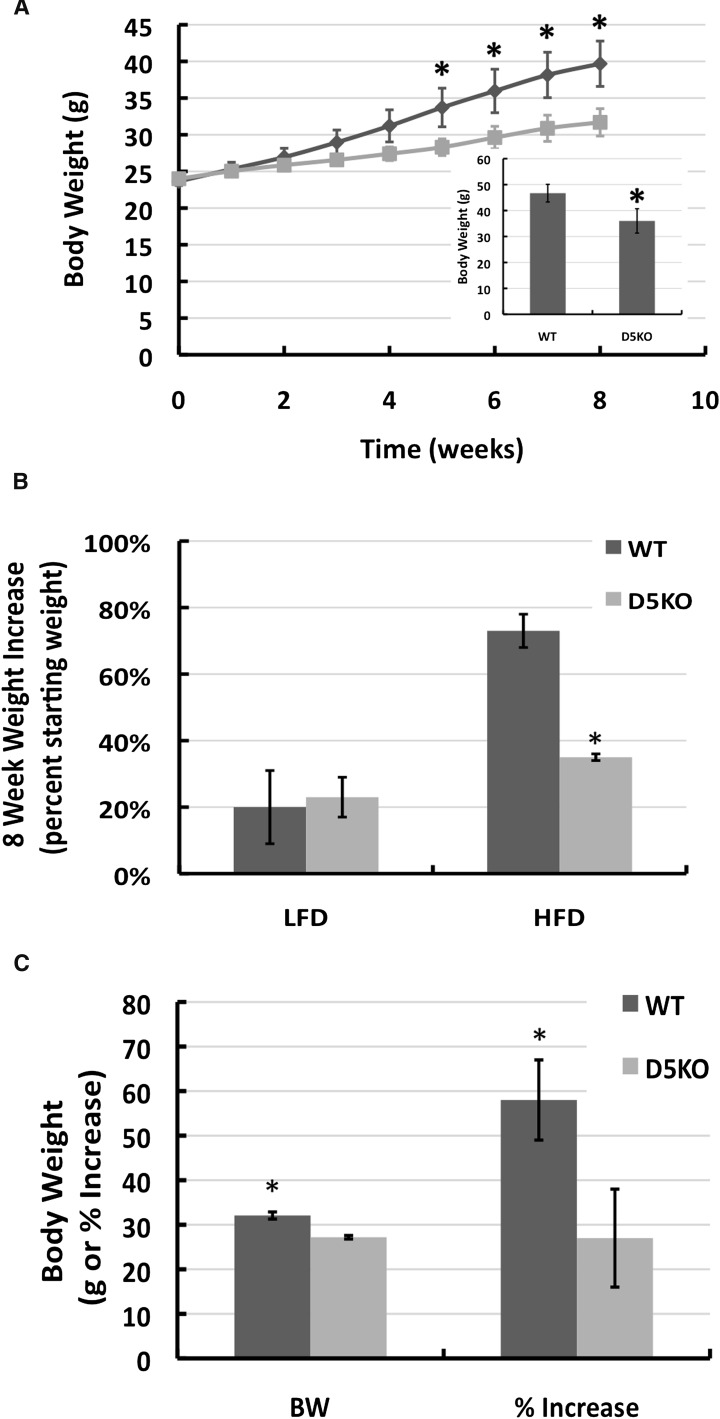
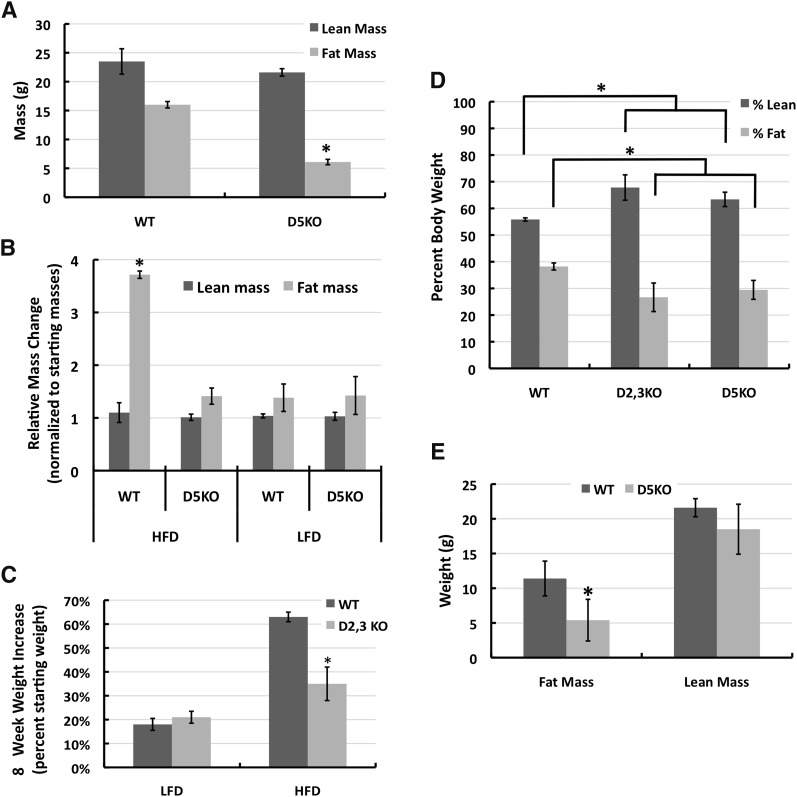
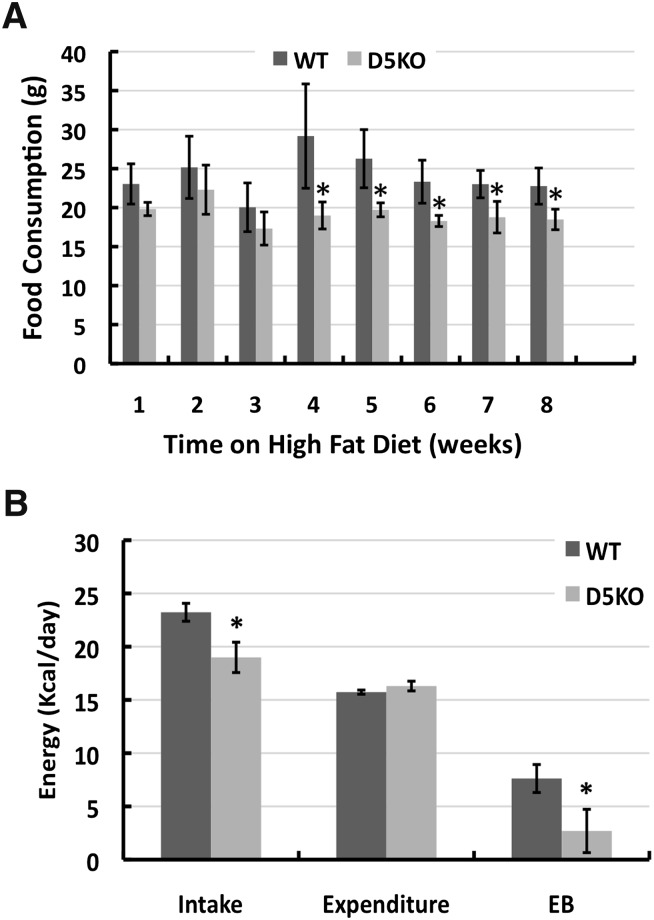
References
-
- Catenacci V. A., Hill J. O., Wyatt H. R. 2009. The obesity epidemic. Clin. Chest Med. 30: 415–444 - PubMed
-
- Unger R. H., Clark G. O., Scherer P. E., Orci L. 2010. Lipid homeostasis, lipotoxicity and the metabolic syndrome. Biochim. Biophys. Acta. 1801: 209–214 - PubMed
-
- Torres D. M., Harrison S. A. 2012. Nonalcoholic steatohepatitis and noncirrhotic hepatocellular carcinoma: fertile soil. Semin. Liver Dis. 32: 30–38 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
