

Two DNA-encoded strategies for increasing expression with opposing effects on promoter dynamics and transcriptional noise
- PMID: 23403035
- PMCID: PMC3668364
- DOI: 10.1101/gr.149096.112
Two DNA-encoded strategies for increasing expression with opposing effects on promoter dynamics and transcriptional noise
Abstract
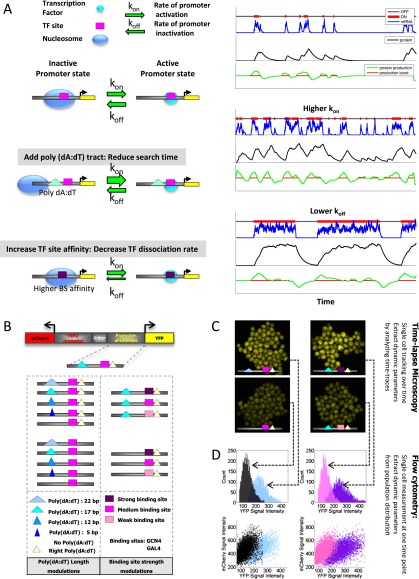
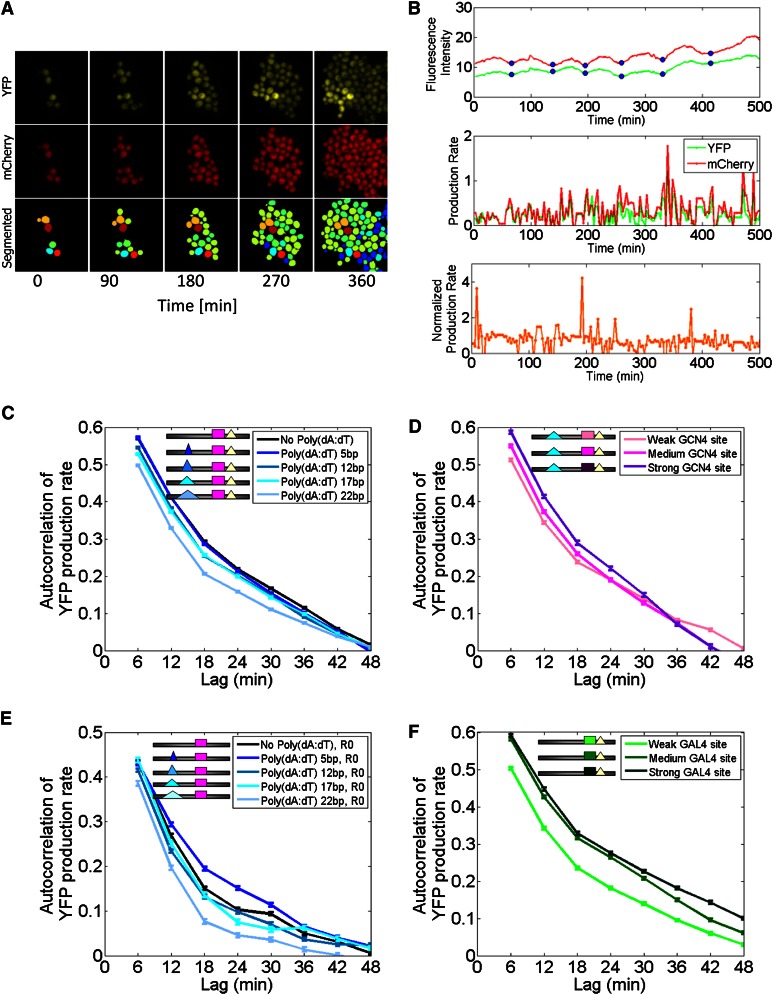
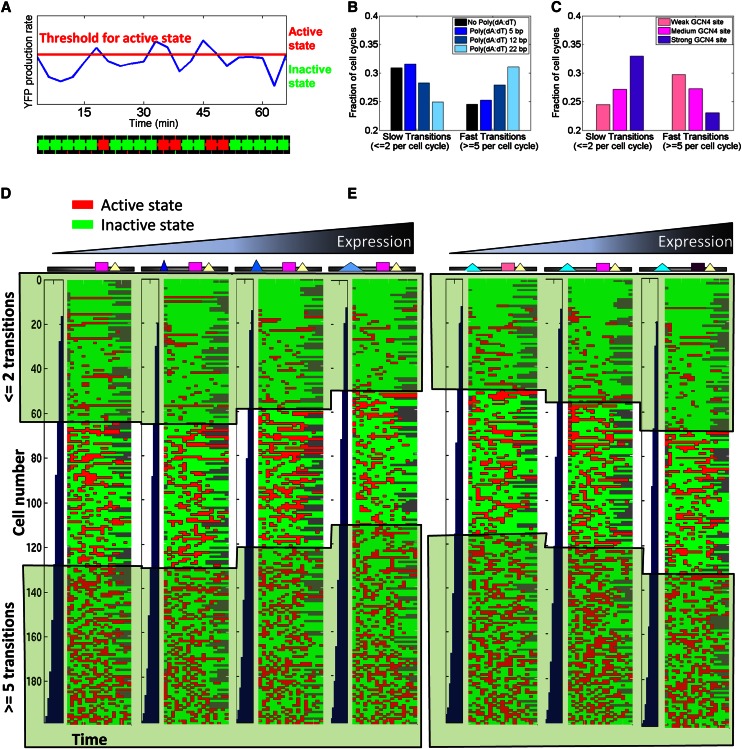
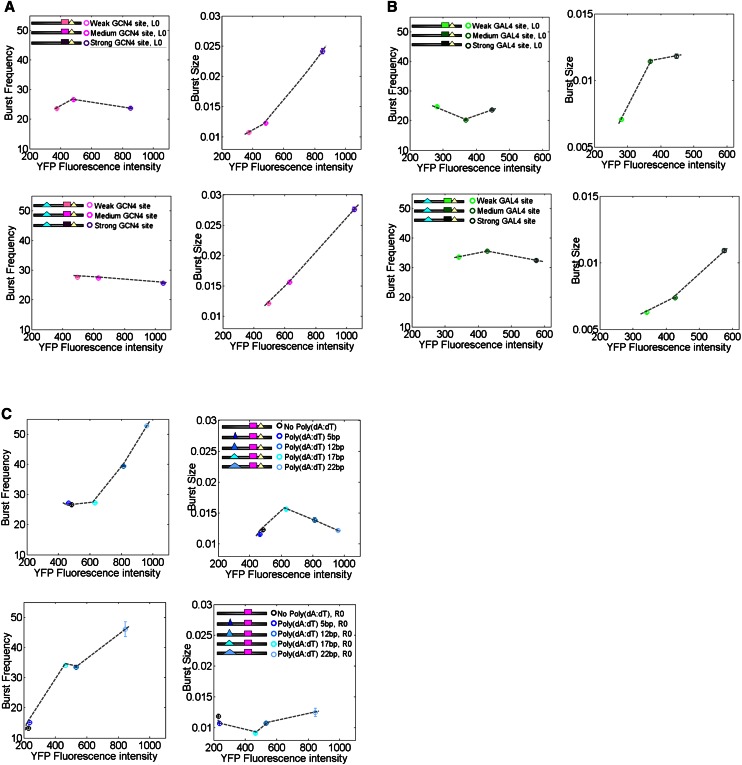
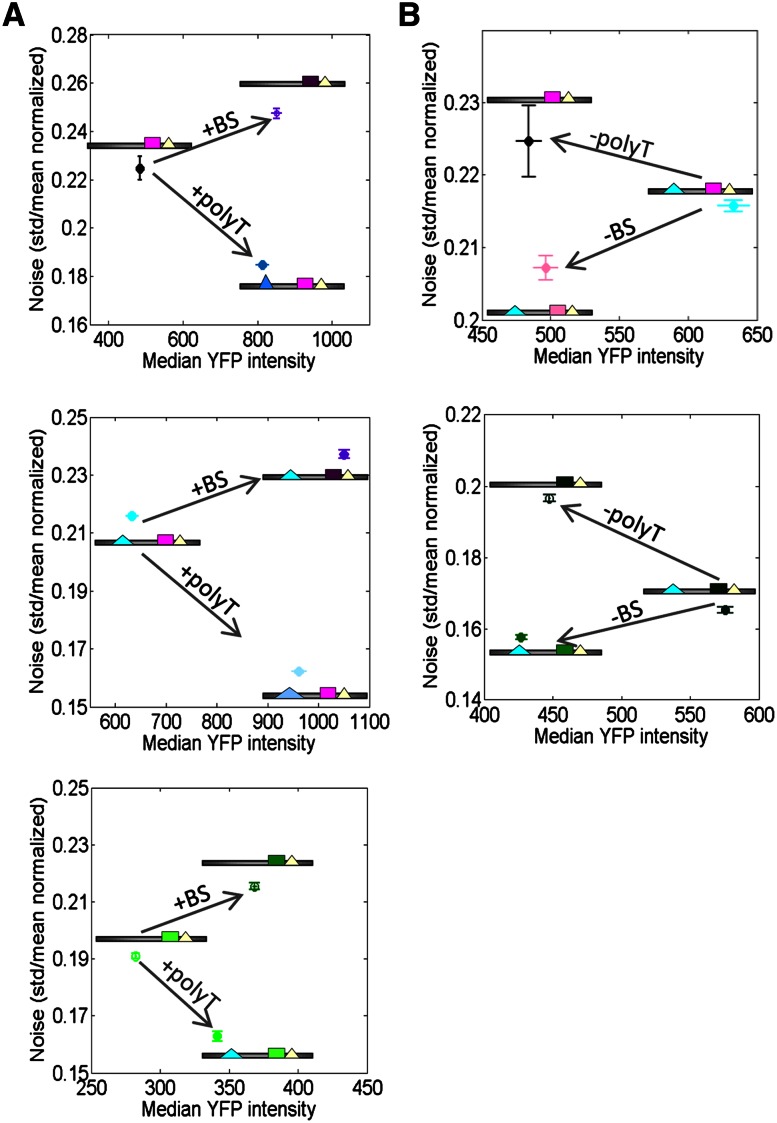
Individual cells from a genetically identical population exhibit substantial variation in gene expression. A significant part of this variation is due to noise in the process of transcription that is intrinsic to each gene, and is determined by factors such as the rate with which the promoter transitions between transcriptionally active and inactive states, and the number of transcripts produced during the active state. However, we have a limited understanding of how the DNA sequence affects such promoter dynamics. Here, we used single-cell time-lapse microscopy to compare the effect on transcriptional dynamics of two distinct types of sequence changes in the promoter that can each increase the mean expression of a cell population by similar amounts but through different mechanisms. We show that increasing expression by strengthening a transcription factor binding site results in slower promoter dynamics and higher noise as compared with increasing expression by adding nucleosome-disfavoring sequences. Our results suggest that when achieving the same mean expression, the strategy of using stronger binding sites results in a larger number of transcripts produced from the active state, whereas the strategy of adding nucleosome-disfavoring sequences results in a higher frequency of promoter transitions between active and inactive states. In the latter strategy, this increased sampling of the active state likely reduces the expression variability of the cell population. Our study thus demonstrates the effect of cis-regulatory elements on expression variability and points to concrete types of sequence changes that may allow partial decoupling of expression level and noise.
Figures
References
-
- Bar-Even A, Paulsson J, Maheshri N, Carmi M, O'Shea E, Pilpel Y, Barkai N 2006. Noise in protein expression scales with natural protein abundance. Nat Genet 38: 636–643 - PubMed
-
- Blake WJ, Kaern M, Cantor CR, Collins JJ 2003. Noise in eukaryotic gene expression. Nature 422: 633–637 - PubMed
-
- Blake WJ, Kohanski MA, Isaacs FJ, Murphy KF, Kuang Y, Cantor CR, Walt DR, Collins JJ 2006. Phenotypic consequences of promoter-mediated transcriptional noise. Mol Cell 24: 853–865 - PubMed
-
- Cai L, Friedman N, Xie XS 2006. Stochastic protein expression in individual cells at the single molecule level. Nature 440: 358–362 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources