

Organization and maintenance of molecular domains in myelinated axons
- PMID: 23404451
- PMCID: PMC4049519
- DOI: 10.1002/jnr.23197
Organization and maintenance of molecular domains in myelinated axons
Abstract
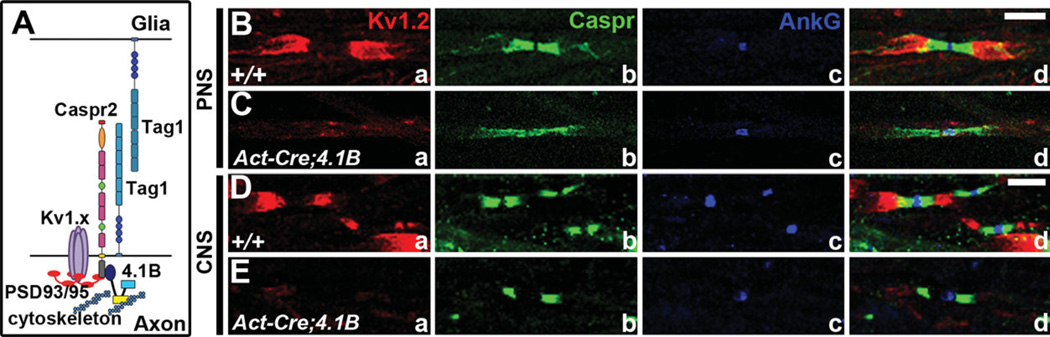
Over a century ago, Ramon y Cajal first proposed the idea of a directionality involved in nerve conduction and neuronal communication. Decades later, it was discovered that myelin, produced by glial cells, insulated axons with periodic breaks where nodes of Ranvier (nodes) form to allow for saltatory conduction. In the peripheral nervous system (PNS), Schwann cells are the glia that can either individually myelinate the axon from one neuron or ensheath axons of many neurons. In the central nervous system (CNS), oligodendrocytes are the glia that myelinate axons from different neurons. Review of more recent studies revealed that this myelination created polarized domains adjacent to the nodes. However, the molecular mechanisms responsible for the organization of axonal domains are only now beginning to be elucidated. The molecular domains in myelinated axons include the axon initial segment (AIS), where various ion channels are clustered and action potentials are initiated; the node, where sodium channels are clustered and action potentials are propagated; the paranode, where myelin loops contact with the axolemma; the juxtaparanode (JXP), where delayed-rectifier potassium channels are clustered; and the internode, where myelin is compactly wrapped. Each domain contains a unique subset of proteins critical for the domain's function. However, the roles of these proteins in axonal domain organization are not fully understood. In this review, we highlight recent advances on the molecular nature and functions of some of the components of each axonal domain and their roles in axonal domain organization and maintenance for proper neuronal communication.
Copyright © 2013 Wiley Periodicals, Inc.
Figures




References
-
- Ango F, di Cristo G, Higashiyama H, Bennett V, Wu P, Huang ZJ. Ankyrin-based subcellular gradient of neurofascin, an immunoglobulin family protein, directs GABAergic innervation at purkinje axon initial segment. Cell. 2004;119:257–272. - PubMed
-
- Arroyo EJ, Xu T, Poliak S, Watson M, Peles E, Scherer SS. Internodal specializations of myelinated axons in the central nervous system. Cell Tissue Res. 2001;305:53–66. - PubMed
-
- Baba H, Akita H, Ishibashi T, Inoue Y, Nakahira K, Ikenaka K. Completion of myelin compaction, but not the attachment of oligodendroglial processes triggers K+ channel clustering. J Neurosci Res. 1999;58:752–764. - PubMed
-
- Banerjee S, Sousa AD, Bhat MA. Organization and function of septate junctions: an evolutionary perspective. Cell Biochem Biophys. 2006;46:65–77. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
